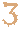| ��ҳ ==> �ܵĹ��� ==> �鿴��Ϣ |
 ������ �����������棬�������Ԫ�����������Ǿ��5.7���������д������ﻯʯ���ֵ�ʱ�ڡ� �������ʾ�����ʱ�ڵ�������������������֡�����Щ���������ߺ��Ѽ��������ʱ��ͳ���������ǰ����͡� ���������һ������Ԫ���档 ʱ���� ��������Cenozoic�� ���ļͣ�Quaternary�� 20����ǰ���ִ�������֡� �½��ͣ�Neogene�� 2300����ǰ~260����ǰ �Ž��ͣ�Paleogene�� 6500��~2300����ǰ ��֪������鳤�ද� ��������Mesozoic�� ���Ѽͣ�Cretaceous�� 1.45��~6500����ǰ ������������鶯��ӹܵ��� ٪�ͣ�Jurassic�� 2��~1.45����ǰ 1.5����ǰ�����ݻ������ࡣ �����ͣ�Triassic�� 2.51��~2����ǰ ����Ŀ���������2.3����ǰ�� ��������Paleozoic�� �����ͣ�Permian�� 2.99��~2.51����ǰ �������� ʯ̿�ͣ�Carboniferous�� 3.59��~2.99����ǰ ���������Ѻ��������档 ����ͣ�Devonian�� 4.16��~3.59����ǰ ���ֵ�һ�ּ����������ưԺ�����������������࣬��ʼ��½����� ־���ͣ�Silurian�� 4.44��~4.16����ǰ ����������α���ٺϣ�ֲ�ﵽ½���������� ���ռͣ�Ordovician�� 4.88��~4.44����ǰ ���dz�Ϊ�����г��������֡� ����ͣ�Cambrian�� 5.42��~4.88����ǰ ��������������ż�Ӳ��ǵ����������Ҷ�档 ��������������Paleozoic������PZ���ǵ���ʱ���е�һ��������ʼ��ͬλ������542��0.3�����꣨Ma����������251��0.4Ma�� ��������������ĵ�һ��������һ������Ԫ�������Ԫ�Ŵ�����һ����������������������������͡����ռ͡�־���͡�����͡�ʯ̿�͡������͡����к���͡����ռ͡�־�����ֺϳ��������������͡�ʯ̿�͡��������ֺϳ����������� ��������ΪԶ�ŵ�����ʱ��������Լ3���ꡣ�Զ������˵������һ����Ҫʱ�ڡ�����һ����������ȫ��������Ľ��������˺���͵���Ļ������Ͷ���Ļ��Χֻ���ں����ڹ�������͢���£���Щ����Ļת������½�ء����������ڣ����ж�������Ʋ��鶯��Ķ�����֣���������������֪����һ���������������ᡣ ���������Ϊ������ʱ���� ����������Ϊ���༰������ʱ���� ���������� 542�C485.4������ǰ ȫʱ��ƽ������O2���� Լ12.5 Vol %��Ϊ�ִ���63% �� ȫʱ��ƽ������CO2���� Լ4500 ppm��Ϊǰ��ҵʱ��16���� ȫʱ��ƽ���ر��¶� Լ15�棨�����ִ�1�棩 ��ƽ�棨�����ִ��� ��30���ȶ�������90�� ����ͣ�Cambrian�����ſڣ���������Ŀ�ʼ�����Լ5��4ǧ1������ǰ��4��8ǧ8�����ꡣ�������������Ӣ������ʿ��һ���Ŵ������������ơ�Cambria�����õصĺ���͵ز㱻�����о�����������Դ�Ծ�ʱ�ձ���ʹ�����ﺺ��������������������͡�������������֥��������֣�kanbuki���� ����Ϳ��ٷ�Ϊ�纮��͡��к���͡�������͡������������������ĵ���ʱ������һ�����ǰ��ռ͡� ��Ҫ ����;��ʮ�־�Զ�����ǶԵ�ʱ�ĵ�ò����һ����֪��ֻ֪��ʱ�Ĵ�½��һ����Ҫ��½�ĺͼ����С����ɡ���ʱ�Ķ��ﶼס�ں������������ů����ƽ�����ߣ���û�˴�Ƭ�ĵ��ݵء�����dz���ش�Ϊ�µ����ֵ��������˼�Ϊ������������������һ�����м�Ӳ�ı��ǻ��ڹ����Ķ�������γɻ�ʯ�����ף���˺���ǰ�����嶯�ﲻͬ������Ͷ��������˴������塣 ����ʹ� ����������ǰ�ķ��ٲ�ʢ����ν�����ݻ�ʷ�ϵĴ�ը���²����Ķ����еĵ��˺����ĩ�Ѿ����������ڲ���˹ҳ�ң�Burgess shale�����״η��ֵĻ�ʯ��������Ҫ�Ķ�����Ⱥ�����ں���ͳ��ֵģ��������������ļ������Chordate���� ����͵Ķ��� ����ʱ���У��������״γ����˴�Ӳ�ǵĶ����Ȼ����͵Ķ����Ǻ�����������Ǻ������δ���������������� ����֪�ĺ���Ͷ��ﶼ�ڼ��ô�IJ���˹ҳ�Ҽ��й���ñ��ɽҳ���з��֡� ����Ⱥ�Ժ���������Ϊ�����ر�����Ҷ�桢��Ϻ���͵�������Ż�������塢����ȿ�ʼ��ʢ�� ����ʹ�����ʹ�����ƺ������������Cambrian Explosion��������Զ�ʱ�ڵĽ����¼�����ʼ�ھ��5.42����ǰ�ĺ����ʱ�ڣ���ʯ��¼��ʾ��������Ķ���š�������һʱ�ڳ����ˡ��������˽������Ĵ�Լ2ǧ����-2.5ǧ���꣬�������˴�����ִ������ŵķ�ɢ�� ����ִ����Ľϸߵ������Լ����ֶ����ԣ����ǣ���һ���α�����س�Ϊ����������Ҳ��������Ŀ�ʼ�� ��������ط��ֵĻ�ʯȺ��ͬӡ֤����һ��������ʷ�ϵ�׳�۾��������ڼ��ô�IJ���˹ҳ�ң������й�����ʡ�ν���ʯ�صȡ���һʱ�ڵĻ�ʯȺ�൱���ͣ��dz���IJ�ͬ��������X��ͬʱ����һʱ�ڳ��֡� ����ʹ�����ʵ֤��Ҳ���ô���ķdz������ڡ�������Դ����д������������鵽����Ϊֹ����û�취���͡����ԣ�������Щ�˸պþͿ����������������������������ݻ��۵㡱������ʹ�����������һ�ٶ���Ľ��գ�����ʹ������ǿ�ѧ���һ�����⣬�д�����Ŀ���֤�ݳ�����Ҳ�����ܿ�����ʱ��ʵ��������ҳ�������ԭ�� ���ܵ�ԭ�� ��Ժ���ʹ���ѧ��������������ּ��裺 1.�����ں����֮��ų������Ա��滯ʯ���ȶ��Ҳ㣬��ǰ����͵ij�������ڵ��Ⱥ�ѹ�����γɻ�ʯ�� 2.���ﵽ�˺���Ͳ��ݻ����ܹ��γɻ�ʯ�ļ�Ӳ���塣 3.�������ۻ��㹻��������������ʹ���������ʱ���ݻ��������γɳ������������⡣ 4.ijЩ��ʳ�Զ������������ȶ�ƽ��ĵ���������ԭ��ռ���Ƶ����֣��ͷ���̬��λ���������֣������ٽ�������������ȵ����ӡ� �������о���Ϊ������ʹ������Ͽ�����ĩ������¼��йأ��ôξ���ȵ�ϸ����������¼������������棬Ϊ��ϸ��������������ķ�չ��ƽ�˵�·�� ����Ͷ��� ��Ϻ ŷ�ͱ���Ы ��ϼ�� �ֵ��� �����dz� ����ͣ����ռ�����¼������-���ռ�����¼���Լ��4.88����ǰ����һ�η��������OȺ����¼�������¼������������棬�����˹������ĺ���ͣ���������ռ͡���ʱ�����˺ܶ����㶯���š�����ʯ�����ؼ�������Ҷ���������Ŀ���ڵ�����Լ49%������������¼�����ʧ��Ŀǰ�����ԭ��û�з��֡� ����¼����� �������� ��ˮȱ�� �������� ����˹ҳ������˹ҳ�ң�Burgess Shale Formation���ڼ��ô�������Ӣ�����ױ��Ǿ��ڵ�����ɽ��������������ѧ�Ҳ��˹�������ء��ֶ����أ�Charles Doolittle Walcott����1909�����ȷ��֡����ص����Ǻ�ɫҳ�ң�ҳ�����г�ǧ�ֻ�ʯ����Щ��ʯȺ�Ա�����������֯������������Ҫ�ֲ����ĺ��ҹ���Yoho National Park���ڵķƶ��£����������������Ļ�ʯ��֮һ�����Լ5.05���꣬�൱�ں�������ڡ��Ҳ���Լ161�ף���Ŀǰ����İ�������������֯�Ļ�ʯȺ֮һ�� ��һ��ҳ��¶ͷ���ڿ�������ҹ��ϲ�42���ﴦ�� ��ʷ������Ҫ�� ����˹ҳ������1909�걻������ѧ�Ҳ��˹�������ء��ֶ����أ�Charles Doolittle Walcott���ڵ���Ұ����齫������ʱ���֡�����1910��ʱ���������������ӻص����أ������ڱ���Ϊ��ʯ�루Fossil Ridge���IJ��潨���˲�ʯ�����ڻ�ʯ���ھ���Ļ�ʯ��Ϊ����������֯�������ã����������������൱�㷺�����ֶ�����֪�������ڹ�����ѧ����Ҫ���·��֣�ֱ��1924����74��Ϊֹ������ÿ�궼�������ھ��ܹ�������65000����ʯ�걾����������˹ҳ�ҵĻ�ʯ���൱���ص������ֶ������ڷ��ֺ�ֱ��1927��ȥ���������ڻ�ʯ�о����ֶ�����ʱ�������Ŀ�ѧ�۵�����ͼ����ʯ���ൽ�ִ����������Ⱥ��ֱ��1962�갢�����С�����������Alberto Simonetta���ٴ����µ�����Щ��ʯ����ʹ�ÿ�ѧ������ʶ�����ֶ����صĹ����������ƿ����ش��ֵ��Ļ������ȷ��ʾ��Щ�����ﲢ������ȷ���ൽ�ִ�����Ⱥ�� ���ֶ����ز�ʯ�����¿��ڻ�ʯ���ж�������Ҷ��ר�ҹ���������Ī������٣�Harry Blackmore Whittington����˵�����ɼ��ô���ʵ���֣�Geological Survey of Canada�����С���һ���ɹ����ѧ�IJ��������ɵ£�Percy Raymond����1924�꽨�����²�ʯ����λ�ڻ�ʯ���ϸ߶�Լ20�״������������λ�������Ľ��Ŵ�ѧ�о�������ˡ������˹�����ɡ�������Ī��˹��ʼ�Բ���˹ҳ�ҽ����������ٵ��飬Ҳ��¶�˲���˹����Ⱥ���ݻ��ֻ����ֶ������о��ɹ������Ҹ����⡣ȷʵ�����ಮ��˹ҳ���ڵĻ�ʯ�����˾��ȵĽṹ���������Һ�������֪����ֻ�������ط����ơ�����ŷ�ͱ���Ы��5���۾���һ������������������ܵĿڱDz���Nectocaris���������Ƽ����ŵ���������һ���������һ��ļ����ֵ��棨Hallucigenia������ĸ�ԭ״̬����ȷ״̬�����෴��������Ϊʹ�����߶ԳƵĴ���·�� �ڼ��ô������ֺ�UNESCOȷ���˲���˹ҳ�ҵ���ʷ��ֵ�Ժ���1970������ڿ�ʼ�������òɼ���ʯ������ѡ���ʯ�ɼ������ɻʼҰ����Բ���ݽ��С��ò���ݵĹݳ���˹�ɵ¡�����˹����������ѧ��ר�ң���ȷ��������������¶ͷ����Щ¶ͷ�ڵز��е�λ�ñ��ֶ����ز�ʯ���ĵز�λ�ø�͡���Щ�ط������Աȿ�ѧ���о�������ٶȷ����µ����֡� ʷ�ٷҡ��Ŷ��µ��顶�������������1989����棬ʹ����˹ҳ���ܵ�һ�����ע�⡣�Ŷ�����Ϊ��ʯ�Ĵ���ֻ���ʾ��ʱ��������ʽ�ֻ����ȱ����ڷָ���������صĺ�����������ݻ����ԡ��Ŷ��¶Ժ���Ͷ���Ⱥ���о��൱�������ɡ�������Ī��˹���ֶ����صij����������о��Ľ������Ī��˹ǿ�ҷ��ԹŶ��µĽ��ۣ�Ī��˹��Ϊ�������еĺ���Ͷ���Ⱥ���Ա��������ִ��������֮�¡� ����״̬ ����˹ҳ�Ҹ�����ʯ�ij�������ʷ�ٷҵز㣨Stephen formation����ء�����ز���һϵ��ϸС�İ�ɫʯ�������Ҳ㣬����Լ5.05���ꣻ�ز�ĵײ���λ��һ����Լ160�����µײ������λ��Զ���ڱ����������ǿ�Һ��ˡ������ֱ�������ǺͿ����ڲ���˹ҳ��֮ǰ��ʱ�����γɵ�Cathedral Formation��ʯ����ɺ��������ɡ��ز����ϸ�γɻ���Ŀǰ�Բ�����������Ϊ���ܵļ�˵����Cathedral Formation��ɺ������Ե��ɺ�����������ַ���������뱻���͵���Զ��ɺ������Ե����������ľ��룬֮��ϲ���Լ5.09����ǰ�ʹ�ز���롣���������һ���ײ��������Ķ������£���ΪCathedral Formation��ʯ���������Ա������ļ�ѹѹ���ġ������ı������ý�һ��������ΪʲôCathedral Formation�����滯ʯ����Ϊ���ѹ���Ҳ������ֱ�и�ʹ��ʯ���ѣ���ɻ�ʯ����ֱ�и�ֶ����ز�ʯ����������Щ���⻯ʯ����Ϊ��Щ��������ʷ�ٷҵز㣬�ò�ʯ��ȷʵ�ھdz��ӽ������ʱ�������±�Ե���� ԭʼ���뷨�в���˹ҳ������ȱ�������г��������������о���ʾ�ڳ���������һֱ���ڡ���ǰ��Ϊȱ��������ֻ�DZ��������������ʹ���ⸯ�ã�Ҳ�ɲ����ʺϱ�����������֯�Ļ�ѧ���������⣬��Ѩ����������١���������Ķ�Ѩ���㼣������������֯��Ȼ�����֣���ȴ��ϡ�еģ���һ����˵��ֱ����ij̶����ޡ���ˮ������һ����˵����μ�����˹ҳ�ұ�����ʽ�� �ز�ѧ ����˹ҳ�ҵز�����10���ز��еĶ���ɵģ������������ֶ����ز�ʯ��ҳ�ҶΣ��öΰ����˴��ģ��Ҷ��㣨Phyllopod bed���� ����ѧ��������� Ŀǰ����������������벮��˹ҳ�����������˹���صز㣨Lagerstatte�������ں���͵������ϱ��������Ҫ�ࡣ������Ϊ�����ھ�Ѩ�Ļ�ڷ�Χ�����ܵ����Ƶġ���Ϊ���������Ŷ���������������Ϳ�ʼ���ձ飬������������֯�Ļ�����ϡ�١���ǰ����͵Ķ��ﻯʯ��¼�൱ϡ���Ҳ����ԡ��� ����Ⱥ ����˹ҳ�ҵ�����Ⱥ�ǵ��͵��к���ͳ�������Ȼ������м�Ӳ��ǵ�����ֻռ��Լ14%������ЩӲ������Ҳ���������Ƶĺ���͵ز��б�����ռ�����Ʊ��������ʾû����������û�м�Ӳ��ǵ���������������ʵ���������������������˹���صز�Ҳ��������Ӳ��ǵ���� ����˹ҳ�ҵ�����Ⱥ�����൱��������������Ӿ���������ϡ�٣���Ҫ�ǵ����������ģ����������ں���Ư����̶��ں�������Լ����֮���IJ���˹ҳ�������ǿ��ź��׳������е��л�����Ϊʳ����������֮һ�������Ǵ�ˮ���˳�ϸС����Ϊʳ����ʳ�Զ����ʳ�Զ�����10%���¡���ʹ��ʳ�Զ����ʳ�Զ�������ϴ��ֲ�ͬʳ�Ե���������ά��һ��ƽ��ı����� ������ʯ��������ʯ�أ��������ν���ʯ�ء������й����ϲ����ϝ����ؾ���40�ദɢ��ĺ���Ͷ����������Ⱥ��ַ�����Լ5.20�ڵ�5.25���꣬��Ҫ������ñ��ɽ��ҳ�ң���ʼ������1984�ꡣ���ڻ�ʯ��ص����������⣬��������������Ӳ�廯ʯ�����ұ����˷dz�������������������ӡ�ۻ�ʯ�������ص��Ƕ�������û�п����ʵ�����֯���ֱ���÷dz��ã������Ƥ����ë���۾�����θ������������ǻ���ȡ����У������������֪����ϵļ������ʯ��5��3ǧ����ǰ����͵ĵز㱻���֡����ƺ����������ǹ����ͷ�Ǽ������� 2012�꣬������ʯ����ѡ���Ϲ��̿�����֯�����Ų���¼����������Ⱥ�ȼ��ô�IJ���˹����Ⱥ�ḻ����������������һǧ���꣬ʹ���ǵ��Կ��������������ʱ���ĺ��ۡ� ���鱣����ʷ �ν���ʯ��������������ʡ��Ϫ�гν��ء��ν��ؾ�����ʡʡ�������еĶ�����60��� 1984��7��1�գ��й���ѧԺ�Ͼ����ʹ������о����о�Ա���ȹ��ڳν���ñ��ɽ������'����'��ʯ���������ʾ��˯��5.3����ĺ�����������硣 1997��5�£�����ʡ������ñ��ɽ�����ܱ�18ƽ������ķ�Χ��Ϊʡ����Ȼ�������� 2001�꣬�ν����ﻯʯȺ����������Ϊ�������ҵ��ʹ��� ���� ��������Ⱥ���°���������� ���ϳ� ���ɺ��� ������� ���Ĵ庣���� ��Ϻ ����� �ų� �ֵ��� ��Ԫ�� ���� ��Ҷ�� ��С��� ���� ���� �˰�ñ����ˮĸ ��Ϻ��Ϻ ��ʯʱ�ڣ� �������������,525�C505 Ma ��ѧ���� �磺 ����� Animalia ��δ�ּ��� ԭ�ڶ��� Protostomia �ţ� ��Ϻ������ Dinocarida �٣� ��Ϻ�� Dinocaridida Ŀ�� �����Ŀ Radiodonta �ƣ� ��Ϻ�� Anomalocaridae ���� ��Ϻ�� Anomalocaris Whiteaves, 1892 �� ���ô���Ϻ A. canadensis (ģʽ��) ������Ϻ A. pennsylvanica ��״��Ϻ A. saron ������˹��Ϻ A. briggsi ��Ϻ��ѧ����Anomalocaris�����룺���Źֵ�Ϻ������һ�����й������������ô��������ĺ���ͳ����Ҿ��з��ֵĹ����������Ϻ�Ƶ���Ϻ�����ǽ�֫�����Ž��ף�Ӣ��������ѧ��Joseph Frederick Whiteaves�״η�����Ϻ��ʯ�����˹�������ء��ֶ����أ�Charles Doolittle Walcott�����ڲ���˹ҳ�ҿ��쵽�������ǵĻ�ʯ����������͢�٣�Harry B. Whittington������𡤲���˹��Derek Briggs����1985��ġ���ѧ���н�������Ϻ�Ļ�ʯƬ������Ϊ��ͬ�����֡� ���� ��Ϻ��һ�ֱ�ƽ���������ζ�������ڽṹ�Ķ������ʹ�����������������֫��ˮ�н��в���״�ƶ�����֫������������ص���һ���γɵ�һ�ġ���Ƭ��������������ˮ���ƶ�����������ĵط�λ�ڵ��������Ը�֮֫�䣬Խ��β���ӽ���Խխ����������ӵ��11�Ը�֫��Խ�ӽ�β�˵ĸ�֫���ѷֱ棬���Ը�֫���������Լ��㡣��Ϻ����һ�Ժܴ���۾������ۣ��������ж��16,000��ˮ���塣 ��Ϻ�ڲ�ǰ��������״��Ϻβ���ĸ�֫����Ϻ�ڲ��ṹ�dz����أ�Ϊ32���ص������������ɣ�����4����������32��С���壬���Ʋ�����Ƭ�����볤��һȦ��Ӳ��������ݡ�����Բ�Σ���ڲ������Ǿ��εģ�������һ�����ݲ����ܹ�����Ӵ�������Ȼ��ˣ�������Ȼ������������С�ͽ�֫��������Ҷ�棬�Լ�������������ҧ������Ϻ����һ�����͵���״β�ͣ��������Ÿ�֫������״�˶����ܿ��ܿ����ƶ���Ϻ��ˮ��ǰ�����������Ƥ�г棨Parapeytoia�������������㡣 ���� ��Ϊ�����õ�Ե�ʣ����Ϲ�����ѧ�Ҳ������Ϻ������ṹ����Ϻ����α����ϡ�������ѧ����Ϊ��Ϻ���첿�븽֫�����������Ӳ�������׳�Ϊ��ʯ��Joseph Frederick Whiteaves��1892����ݶ��ѵĸ�֫��ʯ����Ϻ��Ϊ��һ�ּ����������Ϊ�������������Ϻ��Ϻ��β�������˹�������ء��ֶ����أ�Charles Doolittle Walcott���״η�����Ϻ���첿��ʯ��������Ϊ����ˮĸ��һ�֡��ֶ�����Ҳ������Ϻ������ʳ��ǰ֫����������Ϊ������������β�ˡ����š�Ī��˹��Simon Conway Morris�������첿������������һ��Ļ�ʯ��������ȴ��Ϊ����һ�ּ�������� ��������͢�ٺ�������ˮĸ�������첿��ʯ��ȷ�����ֻ�ʯ������ˮĸTwo large 'arms' (up to seven inches in length when extended���������ֻ�ʯ����ʽ����Ϊ��Ϻ����������ʹ�ù������������������Ƹʳ棨Laggania����Parapeytoia�� ����������ѧ��ʷ�ٷҡ��Ŷ�����Ϊ��Ϻ�ķ��ֱ�ʾ���������ͬ�Ķ������ں����ʱ�����������۵���Ȼ�ܵ�����������ѧ�����ۡ� ��2011��ʱ���������ۻ�ʯ�ڰĴ����ǿ���³�������壨Emu Bay����������ѧ�ҷ��֡�������ѧ����Ϊ������Ϻ�ĸ��ۻ�ʯ�״α����֣�֤ʵ��Ϻȷʵ�ǽ�֫��������Ҳָ����֫����ĸ��ۿ��ԡ��ֱ�з����ѷ�������������������������ʾ���۽������磬�ȼ�Ӳ�����������֫��ӵ��ݻ����硣������ѧ�ҳ�����������Ϊ��Ҷ��ĸ����ǵ���������������ģ������������ʾ��Ϻ�ĸ��۱���Ҷ�滹Ҫ������ʮ��������Ҷ��ֻ�ֱܷ�����ҹ������Ϻ�ĸ��۳�3���֣�ˮ��������֮��ֻ���ִ����ѣ�ӵ��28,000��ˮ���壩�������Ტ�ۡ� ��̬ ��Ϻ�ں���ͺ����зֲ�ʮ�ֹ㷺��������ض����Լ������ǵ��ټ��������й������ô�������������Ĵ����ǣ�����ʱ��Ӻ�������������ڡ���Ϻ��Ҫ�Լ�Ӳ��ǵ�����Ϊʳ��������Ҷ�档��Ϻӵ��ǿ׳���첿�ṹ�����Դ��������ǣ���ʾ������һ����ʳ�ߡ�������ѧ����һЩ��Ҷ�滯ʯ�϶����ҵ�Բ�λ�W�͵�ҧ�ۣ�����ȷ������Ϻ����ɵġ� �ʯ�ṩ������ѧ�Ҹ�����֤�ݣ�֤ʵ��Ϻȷʵ���Բ���Ҷ�档�ʯ�а�������Ҷ����Ƭ�൱����Ϻ��Ψһһ�ֿ�����������������������Ϻȱ��������Ҷ���Ӳ��ǵ����٣����������ƺ����Բ���Ҷ�档������ѧ����Ϊ��Ϻ��������֫�彫������Ҷ�滥��ĥ����˳������Ҷ��Ľ��ʲ�����������ü�Ӳ������ѡ�������ѧ����Ϊ������Ϊ��ʹ��Ҷ�潫������������ⱻץס������ģ����ʾ��Ϻ���첿������ʺ���ʳ������С�͵������������Ҷ����Ƭ�ء� ŷ�ͱ���Ыŷ�ͱ���Ы ��ʯʱ�ڣ� �к���� ��ѧ���� �磺 ����� Animalia ���ţ� ��Ƥ�������� Ecdysozoa �ţ� Ҷ�㶯���� Lobopodia �٣� ��Ϻ����� Dinocarida Ŀ�� �����Ŀ Radiodonta ���� ŷ�ͱ���Ы�� Opabinia ������ Opabinia regalis Walcott, 1912 ŷ�ͱ���Ы��ѧ����Opabinia������һ�ִ���ں���͵������������ֻ��һ���֡���һ��ѧ��ΪMyoscolex��ͬһ��������п����������Ե��ϵ�� ����������л��ڽṹ������δ��������������ܳ���Ϊ40��70���ס�����ͷ������������۱�֧�Ų�ͻ�����۾���������Χ�ܿ��ܵ���360�㡣����ͷ���·����г�����ͬ����һ����Dz����ڳ��ǵ�ĩ��Ϊ��ץ���ԵĴ�״������ֽṹ���ձ���Ϊ��������� �����и��ֽ��������ﶼ��ͬ�������Լ��ܶ���ͬ����һ��ĸ�֫������Ϻ������Ϊ����֮������ṹ������(ͬ�������)������������������״��֫һͬ�γ�β�������ֽ�������֪��ͬ������ڵĽ�֫���ﲻͬ���ǣ�����ͷ���ƺ���û�к����������ڷֿ����� ŷ�ͱ���Ы���ձ���Ϊ��ס��dz�㺣�����������ϣ���������Ӧ����һ�־�����Ӿ�����IJ����ߡ����Dz�������Ϊ������������Ѩ�ڵ�С���ã��������п��������Ծ�������ɳ��������ʳ�������Щ��������ȷ�ģ�ŷ�ͱ���Ы���п��ܾ���������۵����Dz����Խ�ʳ���͵�λ��ͷ���µĿڡ� ������ʷ ŷ�ͱ���Ы���������������ڹ�ȥһ�����������γ��Խ��еķ��ࡣ������˵���������������������ֶ�����������Ϊ���Ǽ��ࡣ ��Ҷ����Ҷ��� ��ʯʱ�ڣ�530�C251.4 Ma �纮��� - �������� ��ѧ���� �磺 ����� Animalia �ţ� ��֫������ Arthropoda �ݻ�֧�� ������ Arachnomorpha ���ţ� ��Ҷ������ Trilobitomorpha �٣� ��Ҷ��� Trilobita Walch, 1771 Ŀ �����Ŀ Agnostida ����������Ŀ Redlichiida �ݰ�ͷ��Ŀ Corynexochida ������Ҷ��Ŀ Lichida ������Ҷ��Ŀ Phacopida ��ͷ��Ŀ Proetida �γ�Ŀ Asaphida ����Ŀ Harpetida �ճ�Ŀ Ptychopariida �ζ�Ŀ Nektaspida ��Ҷ��٣�ѧ����Trilobita�������ͨ����Ҷ�棬�ǽ�֫���������Ѿ������һ�١�������������ں���ͣ��ڹ��������ڴﵽ���壬�˺��������������������Ҷ���ڶ�����ǧ����ǰ�����ͽ���ʱ�����OȺ�������ʧ����Ҷ���Ƿdz�֪���Ļ�ʯ�����֪���ȿ��ܽ����ڿ����������еĻ�ʯ��������Ҷ����������ḻ�ģ������Ѿ�ȷ�����оţ�����ʮ����Ŀ��һ����ǧ������֡��������Ҷ���DZȽϼġ�С�ĺ�����������ں������У�ͨ��������ɳ����ȡӪ������������ֽڣ��д����������Ϊ������ֱ��Ҷ����������ض��з��ֹ��仯ʯ�� ��̬ ��Ҷ��������������Σ�tagmata����ͷ���ɿ�ǰ���������Ϳں���ĸ�����ȫ�ں���һ����ɣ��ز��ɿ�����˶��Ļ���ɣ�β���������β�Ⱥ���ȫ�ں���һ��Ļ���ɡ���ԭʼ����Ҷ��β���Ļ������൱����Ҷ����ز��dz�������ʯ����Ҷ����ʱ�����ֽ�ĵر�һ�����Խ��������һ���������Լ��� ��Ҷ����һ�Կ�ǰ�Ĵ��ǣ�����������֮��û������ÿ�����������ڣ������������ڵĽ�֫�������ơ���һ�ڻ�������ë�Ƶĸ�Ҷ��������������Ӿ���������д���Ҷ����IJ�Ҷ������������Ҷ�ṹ����Ҷ�����ֵ���Դ���������������Ϊͷ���ء�β�����֡� ��Ȼ��Ҷ��ֻ�ڱ����п��ף��������ǵ�����������൱�صģ����ǵ���������ɼ���Ϊ���ĵ��������Ϸ���ʯ�����Ƶȿ�����ɵġ�����������֫���������ܹ�����Ƥǰ��������������еĴֿ����Ҷ������Ƥ�ǽ����п����еĿ���ȫ�����������һ����Ҷ��������¶�����õؿ��ﻯ������������������Ҷ�滯ʯ������������Ƥʱ�����������ͷ�����ز�֮��ֿ�������Ϊʲô������Ҷ��Ļ�ʯ����ȱ��ͷ������ȱ���ز�����ʵ���ʯ����Ҷ���ɵ���Ƥ����������ȥ����Ҷ���γɵġ��������Ҷ���ͷ���������沿���������Ƥ���̡�ͷ����������һ�Ը��ۣ���Щ�ֵĸ����൱�Ƚ�����ʵ��Լ5.43����ǰ��Ҷ���ǵ�һ���������������۾��Ķ��������Ϊ�۾��ij����Ǻ�����������ĵ���ԭ�� �Ӱ��ռ͵������ĩ��һЩ��Ҷ�棨����������Ҷ��Ŀ���������˷dz�����ļ��̣�spines���ƵĽṹ��������Ħ��緢���������Ļ�ʯ������Ҫ���ĵ��������Ħ�����۵Ĵ��м��̽ṹ����Ҷ�滯ʯʵ������α��Ʒ�������ڶ���˹�����������������������Լ����ô���ʡҲ�д����̽ṹ�Ļ�ʯ�����֡����ּ��̽ṹ�����Ƕ�����ij��ֵ�һ�ֵֿ���Ӧ�� �ݡ��¿�ѧ�ҡ���2005��5�£��ı�������Щ������Ҷ������ͷ���������ִ��׳�Ľǡ���������Щ�ǵ���״����С��λ���ش�ѧ�����ʺ�ѧԺ���������ζ���Rob Knell��������Ȼ��ʷ����ݵ�����¡�����ó�������Ϊ����������ΪѰ����żʱ���нǶ����������������ȷ�Ļ�����Ҷ���ǽ���ʷ��������������Ϊ�Ķ�� ��Ҷ��Ĵ�С��1������72����֮�䣬���͵Ĵ�С��2��7���䡣������Ҷ������ȳƳ棨Isotelus rex����1998���ڼ��ô����ѷ����ϰ��ռ͵���ʯ��ֵġ� �о����� ������Ҷ�����۾������ǻ��п���������ζ����������ٵĴ��ǡ���Щ��Ҷ����Ϲ�ģ��������Ǿ�ס�ڷdz���ĺ��ף�����û�й⣬����ò����۾�����Щ���������ξ��۳�Phacops rana���кܴ���۾��� ��Ҷ����۾����ɷ���ʯ��̼��ƣ�CaCO3����ɵġ����ķ���ʯ�����ģ���Щ��Ҷ��ʹ�õ����ġ����ķ���ʯ�������ÿֻ�۾���������������������֫���ﲻͬ���������֫����ʹ�������ġ��ɼ�����ɵ��۾�����Ҷ���̵ķ���ʯ�������˵�����״����۾����������ڽ��ࡣ������Щ��Ҷ��ķ���ʯ���һ���ڲ��ġ����Ͻṹ������ṹ���Խ�����ͬʱ�ṩ���õľ���ڽ�������Ķ�������β����Ophiocoma wendtiiʹ�����Ƶ����� ���͵���Ҷ���۾��Ǹ��ۣ�ÿ��������һ���������⾵��ÿֻ�����ڵ��������ȣ���Щֻ��һ������Щ�ɴ���ǧ���������ĸ���������һ������Ϊ�����Ρ� ���� �����з����������׳汻��Ϊԭ���ڣ�protaspid����������������л�ȫ���ں���һ���γ�һ����һ�Ŀ��ס��ڴ˺������������ÿ����Ƥʱ��β��ǰ�������µ��ز������˺�����Ƥʱ������Ŀ�������ӡ�����Ҷ����׳�����ǵ���ʶ�ܷḻ������Ϊ�о���Ҷ��֮�����Ե��ϵ�ṩ�˷dz���Ҫ�İ����� ���� ��������ͬ��������ʱ��ͷ����ͬ��λ�Ĵ��ڡ���С����״�ر���Ҫ�� ͼ1��ʾͷ���Ĵ�Լ��̬���沿��ͷ����glabella�����ߵIJ�Ҷ����Ҷ����Ƥ�����������գ�librigena��������ֿ�����¶��ͷ�ǣ�cranidium����ͼ2��ʾ��ͷ����ϸ�µĽṹ�� ��Դ ������̬�ϵ�������Ҷ������ȿ����������ڽ�֫����Ķ�����˹������������������氣�Ͽ�����ʱ��������Ҷ��Ķ��������Ҷ���벮��˹ҳ�Һ���������͵Ľ�֫���ﻯʯ���������Ƶĵط��������Ҷ����������֫��������ڰ��Ͽ����ͺͺ���͵Ľ���֮ǰ�й�ͬ�����ȡ� ��� ��Ҷ������ľ���ԭ����������־���ͺ������ʱ�������ǿ��ؽ����ӵ������������������ij�����ͬʱ��������Ҷ�������ļ����ƺ������صġ���Ҷ�����Ϊ��Щ�¶����ṩ�˷ḻ��ʳ� ��������ͺ���ʱ��Ҷ��������������Ѿ��൱���ˣ���ʣ����ͷ��Ŀ��Proetus)��������Ϊ�����ڶ�����-����������¼�������ṩ����������ǰ�İ��ռ�-־��������¼���Ȼû�к����Ķ�����-����������¼���ô���أ�����Ҳ�Ѿ����ؼ�������Ҷ��Ķ����ԡ� ������ڵ�����Ҷ����ӽ��Ķ����������ͷϺ�٣�Cephalocarida���ȶ�� ��ʯ�ֲ� ���������ʿ���ڡ��ر�ż̸��һ�顶̸�ա�ƪ���״μ�����̩ɽ����ʯ������Ҷ�滯ʯ��������һС��һ�ϣ���������٣����ߡ����ߡ�����������������Ȼ��С����������ˮ���·�������ī������Ϊ�⣬��Ի�����ร�⡯����������Ҷ������������������Ļ�ʯһ�𱻷��֣�������ǿ���ȫ���ں����������Զ�ź�������Ҷ����������dz������dz��㡣ż����Ҷ���ں�������ʱ���µ��㼣Ҳ����ʯ���ˡ����������н���Ĵ�½�Ͼ�����Ҷ��Ļ�ʯ�����֣������ƺ�������Զ�ź����о������档 ������ȫ���緢�ֵ���Ҷ�滯ʯ���Է������֣�������Ҷ��ķ�չ�dz��죬������Ƿdz��ʺϱ���������ʯ������ѧ�ҿ���ʹ��������ȷ��������Ҷ���ʯͷ���������Ҷ��������ġ���ù㷺�������Ļ�ʯ������Ϊֹÿ�껹���µ����ֱ����֡�һЩӡ�ڰ��˲�����ʶ����Ҷ����ˮ��������dz���Ҷ��Ϊ��ʯͷ���Сˮ�桱�� ��Ҷ���������������ݣ�Isotelus������˹�����ݣ�Calymene���ͱ�Ϧ�������ݣ�Phacops rana�����ݻ�ʯ�� ��Ӣ�����ױ��ǡ�ŦԼ�ݡ��й����¹�������һЩ�ط����ֹ��dz�ϡ�еġ������������岿λ���㡢���ʹ��ǵ���Ҷ�滯ʯ�� �ڶ���˹���¹���Ħ��硢�����ͼ��ô������ҵ�ɼ���Ҷ�滯ʯ����ҵ�� 2000���ڼ��ô������ʯ��Ҷ��Isotelus rex����720���ף���400���ף�ʱ��Լ4��4ǧ5������ǰ���ռ�ʱ�ڵġ� ���� ����������Ŀ(Redlichiida)���纮���~�к���ͣ���Ϊ��ԭʼ����Ҷ�棬����Ϊ����Ҷ�������Ŀ�����ȣ�Ҳ��Ŀǰ��֪��������ڻ�ʯ�ϵĽ�֫���ӵ��ϸС��β�ͺͽϱ�ƽ���������Լ���İ�����ȫɫ�ۡ� �����Ŀ(Agnostida)���纮���~�����ռͣ��������С��β����ͷ������״��С��ȣ���û���۾������ܺ����ݻ��ɼ��ࡣ �ճ�Ŀ(Ptychopariida)���纮���~�����ռͣ���������������Ŀ�ݻ����ɣ���������Ҷ�������൱�ķ��࣬������ݻ���������Ҷ��Ŀ��ӵ�нϴ���ز���ͨ����β����Ҫ���� �γ�Ŀ(Asaphida)���纮���~��־���ͣ������ճ�Ŀ�ݻ����ɡ����ͽϴ�ͷ����β��ͨ����С���������Ʒ�����ͱ�ƽ������ǿ׳��β���� �ݰ�ͷ��Ŀ(Corynexochida)���纮���~������ͣ������ճ�Ŀ�ݻ����ɣ��Ƿֲ���Χ����㷺����Ҷ����𣬸�������������� ������Ҷ��Ŀ(Lichida)���к����~������ͣ���������Ҷ��δ��ȷʵ���������ϡ���������С�̡�С����Ҫ�����������壬����Ϯ�������Ҿ���ƽ������á����˽�������Ҷ��Ŀ���߳��(Odontopleurida)������������ɳ��߳�Ŀ(Odontopleurida)�� ������Ҷ��Ŀ(Phacopida)������ռ�~������ͣ���������Ҷ��δ��ȷʵ���������ϡ����з���ĸ��۩� ��ɫ�ۣ��Ի����к�ǿ����Ӧ����������з���ľ������١�����Ⱥ�������־����Ҷ�深ʢ����ʱ�����սᡣ ��ͷ��Ŀ(Proetida)���к����~�������ͣ�������ͺ��ڵĴ������Ψһ�Ҵ����Ҷ���࣬���������͵����壬������û�д̺�װ�Ρ� ����Ŀ(Harpetida)���к����~������ͣ���������Ҷ�����ǰ�(rostral plate)����������(facial suture)����ʽʮ�����⣬��һ������ͷ����Ե�ķ������ʽ�� �ζ�Ŀ(Nektaspida)���纮���~��־���ͣ����������桢��С���ȡ� �����Ŀ�����Ŀ ��ʯʱ�ڣ� �纮����� �����ռ� ��ѧ���� �磺 ����� Animalia �ţ� ��֫������ Arthropoda �٣� ��Ҷ��� Trilobita Ŀ�� �����Ŀ Agnostida Salter, 1864 ��Ŀ �������Ŀ Agnostina ���̳���Ŀ Eodiscina �����Ŀ��Agnostida��Ϊ��Ҷ�������һ��Ŀ�����纮��ͳ��֣���������ڿ�ʼ��ʢ�����ռ���������������Ŀ�������Agnostina�� Eodiscina������Ŀ��Ȼ���ָ��Ա��ֳɼ����ơ� �����Ŀ��Ҷ���β����ͷ������״��С��ȣ�ʹ�÷ֱ����ǵ�ͷβ�Ƿdz����ѵģ������������ؽڣ������С��һ�㳤1��3���ף����ٴﵽ5���ף��ֵ������Ŀ��Ҷ����û���۾��ġ� ���ڻ�����������Ŀ��������Ҷ����ݻ��а�����ʲô���Ľ�ɫ�����������Ŀ��������Ҷ���࣬�ֻ��Dz�������Ҷ���࣬���Ǿ�������ġ�������Ϊ�����Ŀ����һ�������׳汻������ӵ�нŵģ��������ǿ��ܺ����ݻ��ɼ��ࡣ ����������Ŀ����������Ŀ ��ʯʱ�ڣ� �纮��� �� �к���� ��ѧ���� �磺 ����� Animalia �ţ� ��֫������ Arthropoda �٣� ��Ҷ��� Trilobita Ŀ�� ����������Ŀ Redlichiida Richter, 1932 ��Ŀ С���γ���Ŀ Olenellina ������������Ŀ Redlichiina ����������Ŀ��Redlichiida����������Ҷ��ٵ���Ҷ�棬Ϊ�Ƚ�ԭʼ�����࣬�����ں�������ڡ�����������ĿҲ��Ŀǰ��֪��������ڻ�ʯ�ϵĽ�֫��� ��������������Ŀ����Ҷ����ͨ��ӵ��һ�����ҳ��ְ�Բ��ͷ������ڵ��ز����С��β��������ͨ��Ҳ��һ������ͻ�������ְ����ε��۾��� �ݰ�ͷ��Ŀ�ݰ�ͷ��Ŀ ��ʯʱ�ڣ� �纮��� �� ������� ��ѧ���� �磺 ����� Animalia �ţ� ��֫������ Arthropoda �٣� ��Ҷ��� Trilobita Ŀ�� �ݰ�ͷ��Ŀ Corynexochida Kobayashi, 1935 ��Ŀ �ݰ�ͷ����Ŀ Corynexochina б�ӳ���Ŀ Illaenina ��dz���Ŀ Leiostegiina �ݰ�ͷ��Ŀ��Corynexochida����������Ҷ��ٵ�����һ��Ŀ�������ں����������͡��ݰ�ͷ��Ŀ��Ҷ����ز�ӵ��2-12�����ڣ����ٳ���12�����ڣ�ͨ��Ϊ7-8�����ڡ� ������Ҷ��Ŀ������Ҷ��Ŀ ��ʯʱ�ڣ� ���ռ� �� ����� ��ѧ���� �磺 ����� Animalia �ţ� ��֫������ Arthropoda �٣� ��Ҷ��� Trilobita Ŀ�� ������Ҷ��Ŀ Lichida Moore, 1959 �ܿ� Dameselloidea Lichoidea Odontopleuroidea ������Ҷ��Ŀ��Lichida���ǵ��Ͷ�̵���Ҷ���ࡣ������Ҷ������ز�ͨ����8-13�����ڣ���һЩ������зdz���Ĵ̡� Ŀǰ������֪������Ҷ��ĿΪ��ӵ�ж�̵��ز�����Ȼ���������������Ϊ�Ƕ����������ij��ֲ�����һ�ֵֿ���Ӧ���������˵����Ȼ����ȫ�Ľ�������������Ϊ���ڰ��ռ�ʱ�ڣ�Ҳ����������������֮ǰ����Щ������Ҷ������Ѿ����������Ƽ���spines���Ľṹ�ˡ� ���˽�������Ҷ��Ŀ���߳��(Odontopleurida)������������ɳ��߳�Ŀ(Odontopleurida)�� ������Ҷ��Ŀ������Ҷ��Ŀ ��ʯʱ�ڣ� ���ռ� �� ����� ��ѧ���� �磺 ����� Animalia �ţ� ��֫������ Arthropoda �٣� ��Ҷ��� Trilobita Ŀ�� ������Ҷ��Ŀ Phacopida Salter, 1864 ��Ŀ ������Ҷ����Ŀ Phacopina ��ͷ��Ҷ����Ŀ Calymenina ��β��Ҷ����Ŀ Cheirurina ������Ҷ��Ŀ��Phacopida��������Ҷ��٣������ڰ��ռͣ�����������������������Ҷ��Ŀ���ֳɾ�����Ҷ����Ŀ��Phacopina������ͷ��Ҷ����Ŀ��Calymenina������β��Ҷ����Ŀ��Cheirurina����������Ŀ����ͷ��Ҷ����Ŀ��Calymenina��Ϊ������Ҷ��Ŀ����ԭʼ�����࣬�������������ճ�Ŀ(Ptychopariida)���ơ� �ճ�Ŀ�ճ�Ŀ ��ѧ���� �磺 ����� Animalia �ţ� ��֫������ Arthropoda �٣� ��Ҷ��� Trilobita Ŀ�� �ճ�Ŀ Ptychopariida Swinnerton, 1915 ��Ŀ �ճ���Ŀ Ptychopariina ���γ���Ŀ Olenina �ճ�Ŀ(Ptychopariida)������Ҷ��٣������ں���ͣ���������Ҷ�������൱�ķ��ࡣ �ճ���ӵ�нϴ���ز���ͨ������8�������Ļ�����ɵģ������ճ�����ز�ͨ���DZ�β����Ҫ���ģ����ǵ�β���DZȽ�С�͵ġ� �γ�Ŀ�γ�Ŀ ��ʯʱ�ڣ� Middle Cambrian�CSilurian ��ѧ���� �磺 ����� Animalia �ţ� ��֫������ Arthropoda �٣� ��Ҷ��� Trilobita Fortey & Chatterton, 1988 �� Anomocaroidea Asaphoidea Dikelokephaloidea Remopleuridoidea Cyclopygoidea Trinucleioidea �γ�Ŀ(Asaphida) ��һ�����ͽϴ߱������Ե���Ҷ��Ŀ. ��ʯ���ֵز�ֲ��ڹ����������������־����֮�䡣�γ�Ŀ��Ҷ���ͷ�� (cephalon) ��β�� (pygidium) ͨ����С������Ҷ���Ʒ�����ͱ�ƽ���ؽ�����ͨ��Ϊ5��12�ڣ�Ȼ��ijЩƷ��ȴ����2���ؽڣ����ж��30�ڵ�Ʒ�֡� ������ѧ���Ʋ⣬���ռ�����ʱ��Ŀǰ����˹ʥ�˵ñ���������Ϊһ��½��������½��֮ˮ�ʻ��ǡ�ʹ��ijЩ�γ�Ŀ��Ҷ���ݻ������۱�����Ƭ�е� Asaphus kowalewski ����һ��ʵ���� �ų��ų� ��ʯʱ�ڣ� ��������ڻ����� ��ѧ���� �磺 ����� Animalia ���ţ� ��ڶ������� Deuterostomia �ţ� �ų涯���� Vetulicolia �٣� �ų�� Vetulicolida Ŀ�� �ų�Ŀ Vetulicolata �ƣ� �ų�� Vetulicolidae ���� �ų��� Vetulicola Hou, 1987 ģʽ�� Ш�ιų� Vetulicola cuneata Hou, 1987 �� Ш�ιų� V. cuneata Hou, 1987 �����ιų� V. rectangulata Luo & Hou, 1999 ��ͷ��ų� V. gantoucunensis Luo et al., 2005 ����ų� V. monile Aldridge et al., 2007 ����ɽ�ų� V. longbaoshanensis Yang et al., 2010 �ų棨ѧ����Vetulicola����������С�ͺ�����ڶ����һ����δ����Ҳ�������ڼ�������������й��ν���ʯ�أ�����ñ��ɽҳ�ҵijν�����Ⱥ���ɽ����Ⱥ��һԱ��ȫ�� 7 �� 9 ���֣������������ͷ��ų棨ȫ��Լ 9.5 ���֣��������������ĵ���˼�ǡ����ϵľ��� ���� �ų汻��Ϊ����������ʳ���������������ܾ�����Ӿ������ǰ����汻��Ӳ�������״�ṹ�����ǣ�������������γ��������ѵ��ѷ죨ÿ�����������ң������о�ϸ����˿���죬������Ϊ�˽�������״�ṹ����������ʹ�����������⡣����ϸ������ǰ��һ�����У��߸�����״�ṹ��ĩ�������ƽ��һ��Ĺų������ͻ������Ш�ιų�Ŀ���ͻ���ҽϴ�����ǰ�����ͽ���Ш�Σ����������ͬ���ģ������ιų������Ҳ���������͡� ����ѧ ���ڹų�ķ����λ������û��ȷ�е�λ�ã��������Ϊ����˫��ǵĽ�֫�����������Ϻ������ȱ���ŵĹ��죬�������ѺͰ�״�ṹ�ȹ�������֪�Ľ�֫��������ǰ��δ���ģ���ˣ��й�������ѧ����¸ɵ����� 2001 �괴�������µķ���ײ㣬��ų涯���š��ų�Ƶȣ�������Щ����ײ�����ں�ڶ������ţ���������������ѧ�� Patricio Dom��nguez Alonso ��Ӣ��������ѧ�� Richard P.S. Jefferies ��Ϊ�ų�Ӧ����β�������һλӢ��������ѧ�� Nicholas J. Butterfield ����Ϊ�ų�Ӧ�����ڽ�֫��� ��Ԫ����Ԫ�� ��ʯʱ�ڣ� ��������� ��ѧ���� �磺 ����� Animalia �ţ� �ų涯���� Vetulicolia ���� ��Ԫ���� Yuyuanozoon Chen et al., 2003 ģʽ�� �����Ԫ�� Yuyuanozoon magnificissimi Chen et al., 2003 ��Ԫ�棨ѧ����Yuyuanozoon�������Ԫ����������Է��ֵسν��Ĺŵ�������Ԫ��������֪�ų��ද����������ߣ��������ǴӴ�����ΰ֮��ġ�magnificent��ת���������ӽ��е�һ����ģ�걾����� CFM00059���������ij���Լ 20.2 ���֡� ���� ǰ������Σ���Ϊ������ڣ��������������ң���ͷ����ÿ����ڽ���һ�����ң�ÿ�����ҿ��ڴ�������˿��������ҡ������Բ���Σ���Ϊ�߸���ڣ���ͬ�ڹų��������������Ԫ���β��ĩ�˲��ʱ�ƽ״��״��������������һֻ�����͵��� �˰�ñ����ˮĸ�˰�ñ����ˮĸ ��ʯʱ�ڣ� 525�C520 Ma ����������� ��ѧ���� �磺 ����� Animalia �ţ� ��ˮĸ������ Ctenophora ���� ñ����ˮĸ�� Maotianoascus Chen et Hou, 1997 �֣� �˰�ñ����ˮĸ M. octonarius ������ Maotianoascus octonarius Chen et Hou, 1997 �˰�ñ����ˮĸ��ѧ����Maotianoascus octonarius���dzν�����Ⱥ�е�һ�ֶ�����������ϳν���ñ��ɽ���ɺ��ȹ��������������������Σ�����Գƣ�Ҳ������Գƣ���λ�������²����룬�����д��ϵ��µİ��о��ȵİ����ΰ��У�������������������ˮĸ�����ţ��١�Ŀ����δ����Ҳ�з���ǻ�������ŵġ��˰�ñ����ˮĸ��������������ˮĸ�����ΰ���Ŀ��������Ҫ�࣬��16���� �������������� ��ʯʱ�ڣ� ����� ��ѧ���� �磺 ����� Animalia �ţ� ��֫������ Arthropoda �٣� Merostomoidea Ŀ�� Limulavida ���� ��������� Sidneyia Walcott, 1911 �֣� ������� S. inexpectans ������ Sidneyia inexpectans Walcott, 1911 ������棨ѧ��Sidneyia����һ��������Ľ�֫����仯ʯ��Ӣ�����ױ��Ǻ���͵IJ���˹ҳ�ҡ� ���� ������泤2-5�����Dz���˹ҳ��������͵Ľ�֫����֮һ�����������ǵ��ܵ���ʳ������ں�����Ѱ��Ӳ���������ʳ���ڵ�����������dz������ݸټ�����ϸС���༰��Ҷ�档 ���˹�������ء��ֶ����أ�Charles Doolittle Walcott����1910���ڲ���˹ҳ�ҿ���ĵ�һ�췢��������档���������ӵ�������������ȡ��Ϊ�䡰���֡��� ��ʱԼ��200���������ı걾�������� �������������� ��ʯʱ�ڣ� �纮��� ��ѧ���� �磺 ����� Animalia �ţ� ���������� Chordata �ܸ٣� ����ܸ� Agnatha Ŀ�� ������Ŀ Myllokunmingiida �ƣ� ������� Myllokunmingiidae ���� �������� Myllokunmingia �֣� ������� M. fengjiaoa ������ Myllokunmingia fengjiaoa Shu, Zhang & Han, 1999 ������㣨ѧ����Myllokunmingia fengjiaoa����һ��ԭʼ��������������ܸٵ����࣬�����ں���͵��й����ϳν���ñ��ɽ������������ֽ��ä���٣���Լ2.8������6���ס� �����������֪����ϵļ������5��3ǧ����ǰ����͵ĵز㱻���֡����ƺ����������ǹ����ͷ�Ǽ����������Ĺ�����δ�г��������biomineralization���ļ��� ��������ģʽ�걾�����й����������������鷢�֡������������Ե�ͷ�������壬�����ı�����������ͷ����5��6���а��������ҡ���������25�ڣ��м�����ʳ������������ĩ�ˡ��ڲ��������Եļ�����������Χ��ǻ��ֻ��һ��������ı걾β�����ڳ������С� ��һ����ͬһ�ز�����������к����㡣Ŀǰ��������ֻ��һ�����֣����Ƿ�����㡣 ������������ ��ʯʱ�ڣ� 535�C510 Ma ��������� ��ѧ���� �磺 ����� Animalia �ţ� ���������� Chordata �ܸ٣� ����ܸ� Agnatha Ŀ�� ������Ŀ Myllokunmingiida �ƣ� ������� Myllokunmingiidae ���� �������� Haikouichthys Luo et al., 1999 �֣� ���Ĵ庣���� H. ercaicunensis ������ Haikouichthys ercaicunensis Luo et al., 1999 �����㣨Haikouichthys����һ��ԭʼ���������������������ܸ١�����ֻ��һ�����֣����Ƕ��Ĵ庣���㣨H. ercaicunensis���� ���Ļ�ʯ�����ϵijν�����Ⱥ��ñ��ɽ�㣩�����֣���Դ�ں���ͣ�����Ϊ�����������ϵ����ࡣ���ķ��ֶԹ�����ѧ������Դ����ѧ˵�м����Ӱ�죬��Ϊ���ѿ�ѧ��ԭ��Ԥ�Ƽ�������ֵ�ʱ��������5����ǰ�� ֧�����ѧ�о���ʾ�����������ԭʼ������ܸ٣���ӽ�����������Լ25���ף�����ͬһλ�㷢�ֵ��������������ģ�걾�����й����������������鷢�֡� ����������ʷĴ��Σ������Ե�ͷ�������ɡ�ͷ���߲������ǣ��ߵ��ǿ��������ң���߶������ǣ�ͷ��������6�������ܶ��9�����������ɷֽڣ���������һ������˫V�ͼ��ڡ������Եı����������м��������DZ걾ֻ������һС�Ρ��߸غ�β������ָ��ͷ�����ֽ�����������������������ġ����������и��������ܳɶԡ��ڸ�����13����״�ṹ����������ֳ�٣������֡��ظ��͡���ֳ��ֻ�����ڸ���ļ��������IJ����ϣ�Բ�����Ѿ����гɶԵ���ֳ�١��������û������ļ��ݲ��ζ�������ǿ����Ϊͬʱ�����ֵ���������ม�Σ�������棬�ɴ˲²⻯ʯ��ͻ���ķ籩������γɡ�������걾�����Դӵز��ϸɰ/��ɰ�Ҳ㣨Լ3�����ϸ����Ҳ㣨�ɴ�50���Ľ��Ӵ����걾����һ��dz�塱�г������з���ѡ���ԣ�������ز�ķ���ƽ�У���б�������Ҳ��еIJ���ͨ���������ڷ�ɰ�Ҳ��еIJ��ֱ���ø��á� ��С�����С��� ��ʯʱ�ڣ� ���������,514�C521 Ma ��ѧ���� �磺 ����� Animalia �ţ� ��֫������ Arthropod �٣� ��Ҷ��� Trilobita Ŀ�� �ζ�Ŀ Nektaspida �ƣ� ����� Naraoiidae ���� ��С����� Misszhouia Chen, Edgecombe & Ramskold, 1997 ģʽ�� ��β��С��� Misszhouia longicaudata ��Zhang & Hou, 1985�� ���� ��β���� Naraoia longicaudata Zhang & Hou 1985 ��С��棨ѧ����Misszhouia����Ϊ���й�����ʡ�ν�ţ�����飨����͵����ף��ز㷢�ֵ�һ������С�ͺ�����֫�����С�ɳ���6���֣���������ƣ�����ֻ��һ�����ֳ�β��С��档 ϰ�� ��С���Ŀǰ����Ϊ��ʳ������ʳ�������֫�Ļ���ʹ���ڽ����ĵط���� ���� ��С�������������п�Ժ�Ͼ����ʹ��������Ĺ�����ѧ���ܹ��٣��Ա������Գν�����Ⱥ�о��Ĺ��ס� ����ʯ����ʯ ��ʯʱ�ڣ� 495�C199.6 Ma ������ͣ��������� ��ѧ���� �磺 ����� Animalia �ţ� ���������� Chordata ���ţ� ���������� Vertebrata �٣� ����ʯ�� Conodonta ��Ⱥ ԭ����ʯĿ Protoconodonta ����ʯĿ Conodonta ������ʯĿ Paraconodonta ������ʯĿ Euconodonta �˵¶���Ŀ Panderodontida ��ݴ�Ŀ Prioniodontida Prioniodinina Ozarkodinina ����ʯ��Conodont���ֳ����δ̣��ɳ����γ桢�����ࡢ�ݡ����ࡣ��һ�ֹŴ�δ֪����IJ������壬��ʼ�ݻ���״������ʯ����С���Ӳ���0.1����4���ף����廯ʯ������Ҫ��ѧ�ɷ�Ϊ����ƣ�һ����������dz�ֻơ��Ұ�ɫ������ʯ�ڸ��ֳ������зֲ����㣬���Һ�ҳ������࣬�����ҡ���ʯ��֮����������ʯ�����ں��������Ǻ����������������δ������ ��������ʯ���������λ����������û�н����20����60�����ǰ����Ҫ������˵�����˵��80�����������������˵�����ֻ�����˵������ʯ����һδ֪�������ڵĹ���Ҳ��ʮ��������ƺ����������õĿ����Ա��ڲ�֧����Ŀ����Դ�һЩ�� ����ʯ�ӹ���������Ϳ�ʼ���֣�����ʢ˥��������������������ĩ�ڣ����ݻ����̴�3���ꡣ����ʯ������ڼ��ݻ�ʮ��Ѹ�٣�ʹ������Ϊ����ʯ����Ч�����ڵز�Ļ��ֺͶԱȡ�����ʯ�����С������̽�����е�������о����м��ͬ�����Բɼ��������Ǵ�ʯ�������ġ������ڵز㻮�ֺͶԱ��У������Ǿ��µز�Ļ��ֶԱ��У�����ʯ������������Ҫ���á� �������� ��ʯʱ�ڣ� ����͵���־���� ��ѧ���� �磺 ����� Animalia �ţ� ��֫������ Arthropod �٣� ��Ҷ��� Trilobita Ŀ�� �ζ�Ŀ Nektaspida �ƣ� ����� Naraoiidae ���� ������ Naraoia ģʽ�� Naraoia compacta Walcott, 1912 �� Naraoia bertiensis Caron, Rudjin & Millken, 2004 Naraoia compacta Walcott, 1912 Naraoia spinifer Walcott, 1931 Naraoia spinosa Zhang & Hou, 1985 Naraoia tianjiangensis Peng, Zhou & Sun, 2012 ���� Naraoia halia ��Naraoia compacta�� Naraoia pammon ��Naraoia compacta�� ���棨ѧ����Naraoia����������������棬��һ�ֲַ��ܹ����������Ҳ���ζ�Ŀ��������������һ�֣��峤Լ1��8���֡� ϰ�� ���е����������������ں��У�����С���һ���������ڶ������г��������Ϊ��ʳ�Ի�ʳ�ԣ���ͷ�����첢���������֮�ϣ��ص��IJ����൱����ʹ������������������ �ֲ� ���滯ʯ�ķ��ִ��ܶ࣬�磺 Naraoia bertiensis �����ڼ��ô���ʡ�ϲ�����������־���͡� Naraoia compacta �����ڼ��ô�IJ���˹ҳ�ҡ������İ�������������ݡ����Ŀ���³���������ں���͵����������ڡ� Naraoia spinifer �����ڼ��ô�IJ���˹ҳ�ң������ں���͵����ڡ� Naraoia spinosa ��������״���棬�������й����ϵijν��أ������ں���͵ĺ��ڡ� Naraoia taijiangensis ������̨�����棬�������й����ݵ�̨���أ������ں���͵����ڡ� �ش����ش��� ��ʯʱ�ڣ� ��������� ��ѧ���� �磺 ����� Animalia �ţ� �ų涯���� Vetulicolia �٣� �ų�� Vetulicolida Ŀ�� �ų�Ŀ Vetulicolata �ƣ� �ش���� Didazoonidae ���� �ش����� Didazoon Shu & Han, 2001 ģʽ�� ���ϵش��� Didazoon haoae Shu & Han, 2001 �ش��ѧ����Didazoon����Ƶش�棬�ǹų涯���ŵ�һ������������������������������������ɽ�Ρ������еġ��ش�Ϊ�й����ʴ�ѧ�ļ�ƣ���С�����DZ����й�����ѧ�Һ�ڱ�����Ը�л����ھ����Ĺ����� ������������ ��ʯʱ�ڣ� ��������� ��ѧ���� �磺 ����� Animalia �ţ� �ų涯���� Vetulicolia �٣� �ų�� Vetulicolida Ŀ�� �ų�Ŀ Vetulicolata �ƣ� ������� Beidazoonidae Shu, 2005 ���� �������� Beidazoon Shu, 2005 ģʽ�� ���������� Beidazoon venustum Shu, 2005 ���� Bullivetula variola Aldridge, et al., 2007 �����ѧ����Beidazoon����Ʊ���������ų棨Bullivetula variola�������ڹų��ද�������ͽ�С��һ�֣�ȫ�� 0.8 �� 1.4 ���֣����������������������������������ɽ�Σ���ɽ���漰���Ĵ����棩�������DZ�����ѧ�ļ�ƣ��������������Ĵ���������˼�����ݻ�ʯ����ü�Ϊ���������� ���� ����С�����ͽ��ƹų棬��������ͻ����ǰ��ͬ���γ��Ŀ飨���Ҹ���Ƭ����Ӳ�İ�Ƭ����ǰ�帲�ǣ�����������Сͻ���������������ң�����Ҳһ����Ϊ�߸���Ӳ����ڡ� ���ռ����ռ� 485.4�C443.4������ǰ ȫʱ��ƽ������O2���� Լ13.5 Vol %��Ϊ�ִ���68% �� ȫʱ��ƽ������CO2���� Լ4200 ppm��Ϊǰ��ҵʱ��15���� ȫʱ��ƽ���ر��¶� Լ16�棨�����ִ�2�棩 ��ƽ�棨�����ִ��� 180�ף������210�ף���ĩ������140�� ���ռͣ�Ӣ�Ordovician������O���ǵ���ʱ����������������ĵڶ����ͣ�Լ��ʼ��4.9����ǰ��������4.38����ǰ�������ա���Ordovices��֮������Ӣ������ʿһ�Ŵ��������˲������ά˹�ˣ�����ѧ�Ҳ��˹��������˼�������������������������Դ�Ծ�ʱ�ձ���ʹ�����ﺺ�������������������ռ͡������������`�ɩ`���������֣���d��ki�������ռͿɷ�Ϊ����������а����������������� ���� ���ռ�����dz���㲼�������ʺϣ������������һ����ʢ�������Ϊ������ռ�������ơ�dz�������ŵ��ܵ���Ҷ�桢�����ࡢ˫���ࡢ�����ࡢ�����ࡢ���ٺϡ������յȡ��峺��dz������̦�桢ɺ���������ࡢ��ࡢ�����ݡ�����ʯ����׳桢����ȡ���ˮ��������ˮ�����ײ��ʺϵ����������棬����Ư���ű�ʯ�ࡢ�б��ǵ���С�����ҶϺ�࣬��Щ����˳����Ư������Ϊ�㲼�Ļ�ʯ�� ��ʯ����������к����������ռʹ�����ʢ����ҪӪƯ����ʽ���ֲ��㣬�ݻ��졢���ױ��棬�ǰ��ռ���Ҫ�ķִ���ʯ������������������α�ʯΪ������������ʯ���е�������Ŀ������ʢ������������Ŀ�ĵ��ʯ���֣��С���������������ʯ��Ķ�ʢʱ�ڡ�������������������������������ռ����ַ��࣬���νϴ���1�ף������ǰ��ռͺ����������͵Ķ����Ҷ���������ܶࡣ���ռͿ�ʼ���ֵĻ������ʢ����Ҷ�����γ��ࡢб�ӳ��ࡢ�����ࡢ�������ࡢ���۳��ࡢ��β���ࡢ��ͷ���ࡢ���߳��ࡢ���߳���ȣ����������нʸٵ����α��ࡢŤ�±���ռ��Ҫ��λ�� ����ϵļ�����������㡢�������Ҳ����ʱ��ʼ���֡� ��ʷ �й����ռ͵ĺ������ں���ͺ������������ġ����ӡ��г���̨������Ե�ش������С�������������������������������ռ����ڵز�ȱʧ�����µİ��ռ͵ز��뺮��ϵ�ʼ����ϽӴ������г���̨���С����������ӵ�̨�����ռ͡�����͵ز�Գ����ϽӴ����а�����֮���г���̨����Ϊ½�أ���������Ե�����⣬��������û�г����� ���ռͼ��ﶫ�˶��ڵ�̨������ΪƵ�������˶����ز����н϶�Ļ�ɽ�緢�ҡ��л��Ժ������Ի�ɽ�ң��籱���ز����� ŷ�Ǵ�½��4���ȶ��ĵ�̨��������˹��̨���������ǵ�̨���г���̨�����ӵ�̨���������������⣬�����ϱ���ˮ���룬�γ�dz��ˮ��̨��Χ���ز���Χ�ơ�����˹�����ӵ�̨����Ե���ʶ���������״�Ĺŵ��к�����Եֹ�ڷ��ޱ������������뵺�в��������ϲ���ӡ�Ȱ뵺���������Ͼ����ϰ뵺��Ĵ����Ƕ����������ռ͵ĺ������������Ͽ������ӵ��ϼ������� ������Ϊ��̨dz������������ʯӢɰ�ҡ�ҳ�ҡ�̼������Ϊ������Ȳ�������½�Ķ�����Ϊ�ز����ĺ�����������м�ҡ�̼������Ϊ����������Ӳɰ�ҡ����Һͻ�ɽ�ҡ���������̫ƽ���ذ��ش�Ϊ�زۺ����б���Ϊ��̨dz������ �ϴ�½����Χ��Ե�ش����ز�������̨�ͺ���Χ�ƣ����ޡ�ӡ�Ȱ뵺���Ĵ��������ϲ��������������ϼ�������Ϊ½�ء� ���� ���ռ��硢���ڼ̳��˺���͵�����������ů�����ֹ㷺�������ϴ�½�����������ģ�Ĵ�½���Ǻͱ�����������������ļ������� ���ŵش����ݣ����ռ��ϼ�λ�����ڱ�����������������ޱ��Ӳ�ķֲ����ϼ�Ȧ�ڵĽ������Ǻϵġ��ϴ�½�Ķ����Դ��ڳ���������������������ǡ��й������������ҳ������Ʋ�Ϊ��������������γ�ȵ��������ռͱ���λ����̫ƽ��½��������λ���ϰ��ӳ��������ʱ�ϰ��������ִ��Ƚ����ԡ�������������ĩ�ڴ���ڵĴ��ڣ�Ӱ��ȫ��ƽ����½���������㷺�ĺ��ˡ� ������������ ��ʯʱ�ڣ� ���ռ�������� ��ѧ���� �磺 ����� Animalia �ţ� ���������� Chordata ���ţ� ���������� Vertebrata �ܸ٣� ����ܸ� Agnatha �٣� ������� Ostracodermi �����㣨ѧ����Ostracoderms����������Ƥ�࣬���������һ��û������ԭʼ���࣬�������й��ʵļװ帲�ǡ����ǵĻ�ʯ�ڱ�����ŷ�İ��ռͼ�����͵ز㶼�з��֡�����һ�����30���ף��������ж�����������ˮ�Ķ�� ������������ֻʹ����������������������ʳ�Ķ����������֮ǰ�����ﶼ��������ͬʱ��������ʳ�ġ����ǵ�ͷ����������������ң��ǿ�����û�����ǡ������㲻�������ļ���������ë������ʳ������Լ�������������ϸС����� ���� ��������Ҫ�ֿ����࣬�н�ԭʼ������Ǹټ�ͷ����١�ͷ����ٱ�����Ǹ���ʤ��������������ȶ�����������Ӿ�� ��Լ��4��������������ܸ٣����ִ����ࣩ���������������½��������������������� ��������Բ�ڸ٣�������Ŀ����ä���٣�һͬ������������ܸ�֮�¡�������������������Ѿ�����ʹ�ã���Ϊ�����Dz�ϵȺ���ԴȺ���������������ǹ������и��мװ弰û����������ķ���ʽ���ơ� ������������ ��ʯʱ�ڣ� 489�C358 Ma ��ѧ���� �磺 ����� Animalia �ǽ磺 ����������ǽ� Eumetazoa �ţ� ��Ƥ������ Echinodermata ���ţ� ���ٺ����� Blastozoa �٣� �����ո� Cystoidea ���������������һ�༬Ƥ����Ǽ�Ƥ����������ϡ�������֯���һ�١���״Ϊ���Σ���ƻ�����ƣ�����������Ϊƻ���Ĺųƣ�������������ಿ�ǰ壨��壩���13��200�飬����Ͼ��а�ף�����ϵͳ���ӡ������ո�Լ��100�����ң��������ڰ��ռͺ�־���ͣ�����������ھ����� ��ʯ����ʯ�� ��ʯʱ�ڣ� 510�C350 Ma �к���ͣ���ʯ̿�� ��ѧ���� �磺 ����� Animalia �ţ� ���������� Hemichordata �٣� ��ʯ�� Graptolithina Ŀ ���α�ʯĿ Dendroidea �ܱ�ʯĿ Tuboidea ǻ��ʯĿ Camaroidea �DZ�ʯĿ Crustoidea ����ʯĿ Stolonoidea ����ʯĿ Graptoloidea Dithecoidea ��ʯ����һ��������ĺ�С��Ⱥ��������������ں����������ʯ̿�����ڡ�־����ʱ�ڵı�ʯ��ʯ���࣬����Ϊ����ʯʱ��������Ϊ�����ƹŴ�����ʹ�õ���ë�ʣ��ʵ�����ʯ�������ڱ�������ǻ�������ѧ���������о�����ȷΪ������� ־����־���� 443.4�C419.2������ǰ ȫʱ��ƽ������O2���� Լ14 Vol %��Ϊ�ִ���70% �� ȫʱ��ƽ������CO2���� Լ4500 ppm��Ϊǰ��ҵʱ��16���� ȫʱ��ƽ���ر��¶� Լ17�棨�����ִ�3�棩 ��ƽ�棨�����ִ��� ��Լ180�ף��ж��ڵ��½� ־���ͣ�Silurian���ǵ���ʱ�����������ĵ������ͣ�Լ��ʼ��4.4����ǰ��������4.1����ǰ�� ־��ϵһ��Դ������ʿ����һ�����ϲ���־���ˡ�1835�꣬Ӣ��R.I.Ī��ѷ��������ʿ���������˹����־��ϵ������ϵ���˻��֣��ñ�ʯ����ʯ���жԱȡ�1879�꣬���˹��������˼��Ī��ѷ�����־��ϵ�²�����Ϊ����ϵ��������Դ�Ծ�ʱ�ձ���ʹ�����ﺺ����������������־���͡������������륭�������֣�siruki���� �ز� ־���͵ز�������ֲ��Ϲ㣬dz���������ǡ�ŷ�����Ĵֵ��������Ĵ����ǵIJ��ֵ��������ޡ��ϼ���Ϊ½�ء� ־���͵ķֲ�ϵͳ����ʯΪ��ѡ���࣬����ά��������ˡ�����3��ͳ����Ӣ��ȷ�������⣬��Ų���ϲ������ô��İ��ٿ�˹�ٵ������ĸ���������ڿ����IJ������ǵ������ݿ˺�˹�工�˵IJ�������������з������õ�־���͵ز㡢����Ⱥ�� ־��ϵ�Ķ��磬��ѡ����һ����ʯ��Monoraptusuniformis��������ĵ�Ϊ�磬ѡ��ݿˡ�˹�工�˵İ����µ�������������Ϊ����߲������档־��ϵ�ĵ磬�ñ�ʯ��Parakidoraptus acuminatus�����ĵ�Ϊ�磬���߲���ѡ���ո���Ī�������ĵ���˹�֣�Dobs Linn�����档���ڸ�������ʹ��츴�ӣ����һ����ʯ���������������������ڵ������������������д��������顣 ־��ϵ���ٷ֣�����4��ͳ�� ����ά��ͳ����������Ӣ������ʿ�ϲ���ѵµ�����ά������Χ����3���ף�³���ס��������ᰲ�ס�������ס� �����ͳ����������Ӣ����ʲ���տ�������˵�������2���ף�����½ס���Ĭ�ס� ����ͳ����������Ӣ����ʲ���տ��������������2���ף���˹�ؽס�¬�¸��½ס� �������ͳ���������ڽݿˡ�˹�工�˵İ����µ����� ���� ־���͵����������ռ������ϵ���У����������ھ������ռ�ĩ����¼�����һ���µĸ��սΡ� ��ʯ�Ǻ���Ư����̬��������עĿ�������ʯ�Ե���ʯ��Ϊ�����絥��ʯ������ʯ�����ʯ���ұ�ʯ�ȡ���ʯ�ֲ��㣬�ݻ��죬�ڵز�Ա����ж��صļ�ֵ�� ��dz�����������У����㶯���������ࡣ�dz��ɳ���80���ף���������̨���巿���������������һ�࣬ʼ��������������־���Ͷ�ʢ�������塢�����塢ʯ���壬���ռ�֮���ȶ���չ��������Ϊ���㶯�����Ҫ��Ⱥ���ڰ��ռʹ�����ֳ�����α���Ť�±�����־��������˥�䡣 ɺ���Ͳ�׳�Ҳ�Ϸ�ʢ�����γ����ク������������㡣����ɺ��������ɺ��������ɺ�������������ล�������������ĺ��У���������࣬�⼸��ɺ������׳�������͵��ﶦʢ��������������� ����ռ���ȣ����������Լ��٣����������Ѹ�������й�������ʢ�����Ĵ���ʯ(Sichunocers)Ϊ���������ݡ����ٺ�������Ϊ��ʢ��һ�ּ�Ƥ������й�����ά��ͳ�г����Ļ��꺣�ٺϣ�Petlocrinus����������״���ִ��IJ��ܴ���Ҷ�浽־��������˥�䣻ֻ�ھֲ��ɼ������й������������ڳ�(Coronocephlus)����������(Eurypterids)��־�����������е�ʳ�������������Ӿ����Ϣ�ں�����ˮ������ˮ�С� ����ʯ�ݻ��졢�ֲ��㣬�̱�ʯ֮�Ա�־���͵ز����һ��Ҫ�Ļ�ʯ��־���ͱ����˿ɿ����������ʯ���������٣��й�־���͵��㻯ʯ��Խ϶࣬����������ล���������ڵز㡣 ��ʷ ־���ͳ��ڣ��ϼ�����Ѹ�����ڣ�����־���ͺ���������������γ������ִ������ԣ��������Խ�ů���������ϵͣ��׳�����������γ�ȵĸ����ɴ�½�⣬���ڸ��ȡ���ů�������¡� ������ά�������ڣ���ƽ����������������ձ��ȱ���������㲼�ĺ�ɫ��ʯҳ�ң� ̼�����Һ����ク�Ĺ㷺�ֲ����ڱ�������ŷ��Ϊ���ԣ���ʾ�˽���ů�Ĺ������ص㣻 ־���͵ĺ���ձ鷢�������й����ϡ��������ǡ�������˹̹�����ĺ��Ȱ���о��з��֣� ־���͵������ҷ������������ǡ��Ĵ����ǵȣ���ӳ�˸ɺ������ȵĹ�������ò�� ���������� 419.2�C358.9������ǰ ȫʱ��ƽ������O2���� Լ15 Vol %��Ϊ�ִ���75% �� ȫʱ��ƽ������CO2���� Լ2200 ppm��Ϊǰ��ҵʱ��8���� ȫʱ��ƽ���ر��¶� Լ18�棨�����ִ�4�棩 ��ƽ�棨�����ִ��� һ���ȶ���189�ף����½���120�� ����ͣ�Ӣ�Devonian������D���ǵ���ʱ���������еĵ��ĸ��ͣ���ʼ��ͬλ������416��2.8�����꣨Ma����������359.2��2.5Ma�� �������Ӣ���н�Devonian����������Ӣ�����Ŀ�����õص�����͵ز㱻�����о����������Եijƺ���Ӣ�Ĵ�ͬС�졣��������Դ�Ծ�ʱ�ձ���ʹ�����ﺺ��������������������͡����������ǩ`�ܥ������֣�d��bonki���� �����������ާ��١����ں��������ɺ��������ԭʼ��ʯ��������֡�����ԭʼ�����ࡢ�Գ�����֣�ާ��ֲ���ԭʼ����ֲ����֣��������������� �����4.05����ǰ��3.65����ǰ�� ��������������������������ͷ�Ϊ������������������������������ ʯ̿��ʯ̿�� 358.9�C299������ǰ ȫʱ��ƽ������O2���� Լ32.5 Vol %��Ϊ�ִ���163% �� ȫʱ��ƽ������CO2���� Լ800 ppm��Ϊǰ��ҵʱ��3���� ȫʱ��ƽ���ر��¶� Լ14�棨�����ִ�0�棩 ��ƽ�棨�����ִ��� ��120�����ִ�ˮƽ����ĩ���ȶ�������80�� ʯ̿���ǵ�����ʷ�е�һ������ʱ��������1822��ʯ̿����Ӣ�����Ѿ���������һ������ʱ���еļ��ˡ�ʯ̿�͵�����������ʯ̿��ʱ����ȫ��������γɵ�ú������3.55����ǰ��ʼ��������2.9����ǰ����������ͺ������֮��ı߽�������Ҫ��ͨ��������ͬλ�ضϴ���õġ� ��ʯ̿�͵��ڲ������������ʹ�÷dz���ͬ��ϵͳ����Щ��ͬ��ϵͳ����ʹ�õ����ʹ�ͳ��ȴ�൱�ȶ�������ŷʯ̿��һ�㱻��Ϊ���������Ǽͣ����������������ȼͺͱ��������Ǽͣ��ڶ���˹�������������Ǽ͡�������ѧ�ϵ�ϸ�ڻ���һ��ʹ�ú������ͷ��ٶ������ٶ��ɺ���ȡ�½��ֲ��Ҳ����������ʯ̿����ϸ�ڻ��֡� ������ ֲ�� ʯ̿�ͣ���������ʯ̿�Ϳ��Ա���Ϊ��ާ��ֲ���ʱ��������ֻҪ����һ�£�����������ľͷ�����ݱ�Ϊһ������ú����ô��ʯ̿��ʱ��ާ��ֲ��ɭ�ֵĹ�ģ�Ϳ��Դӽ���ʯú��Ĺ�ģ�п������ˡ���Щ��Ϊ�����ʯú��ֲ��������Ҫ������ľĿ�ͷ�ӡľ����ֲ���Щ��״��ֲ�����ڽ����ʯ�ɸ٣����ǿ��Դﵽ40�ߣ��侥��ֱ���ɴ�1�ס������ʾ���«ľҲ�ﵽ��20�ߡ������Ŀֲ��Ҳ��Ϊ��һ��ĸߴ�ľ��ֲ�ʯ̿��ĩ���л�ֲ�����ֲ������ɼ������ɼ�����֡� ���� �������� ����ͺ���ռ֧�����õĴ���Ӳ��װ����������ͺ�������¼���û���ٻָ�������ʯ̿�ͺ��е���Ҫ�����ǻ���ķ������ࡣ ���ٺ���ʯ̿�ͳ��ֵ�������������ڼ�Ƥ�����Щ������ʯ̿�͵ĺ����γɲݵ�һ��Ĵ���������棬�������������ʯ�������������ʯ�Ķ�����̦�����ŵĶ����ϣ�߸��ѺͷĴ��ѣ��������ǵ�ϸ����������ǿ��Դﵽ10���״� ½�ض��� ������������������������ʱ���ͳ����ˣ�����ʯ̿���������������档��Щ���滹���۵����ǵij�������ѵȣ���ʯ̿ͳúϵ�ز��з��ֳ���500�ֵ����档ʯ̿��ʱ��½���������Ψһ�ļ����������ܶ�������ǻ��������൱��ˮ��ϰ�ԡ�����������½���ϻ�û�о������֣�������ǵ�����dz��࣬��Щһֱ��6�׳��� ����ʯ̿�͵�ĩ�ڻ��ҵ�������ϵĿ����������ж���Ĺ�����ʯ����ʱҲ����������Ĵ���Ӳ�ǵĵ��� �ŵ��� ������ ��������б����ؿ�ͱ�ŷ-����˹�ؿ��ϵ�һ������½������ĸ����Ǵ�½���������֣�����ķ��ޡ������ޡ��ϼ��ޡ��Ĵ����Ǻ�ӡ�ȣ�֮�䲿�����ɲ�ͬ�ĵ�����ɵĺ��������������Щ�����뱱��-��ŷ-����˹��ɵĴ�½�ѿ�ʼ�нӴ��� ʯ̿�͵ķ�չ ʯ̿����������̼�����չ�����ϡ���ʯ̿�ͽ����ʱ��������̴ﵽһ���߳���ֱ����ʯ̿�ͷ����������뱱��֮��Ŀ�϶�ű��������������ɽ������ɽ�˶���ɡ��������ڽ�����������������˹��ӣ�������ɽ���γɣ��̹Ŵ�½�γɡ� ���� ʯ̿�Ϳ�ʼʱ�����Ͻ�λ�ڵ�����ϼ���ʯ̿���и����Ǵ�½��˳ʱ�ӷ�����ת����������ʱ�ϼ���λ���ϼ���ʯ̿�Ϳ�ʼ�������½�������ʯ̿�;��Ѿ��б����γɣ�����ʯ̿��/�����ͼ��ڱ�����չ���˸߳��ڡ��ڸ����Ǵ�½�����������ҵ������ĺۼ������ʷ���֤����ʯ̿�������±Ƚ���ů��ʱ�������±ȽϺ����ʱ�ڲ��Ͻ��档��ʯ̿�͵Ĵ���ú�ij��������뺣��IJ������²���������������������ڸ����Ǵ�½�ϲ��������ڻ����ӳ�����ɵġ� ����������������������Ӣ�Mississippian���ǵ�����ʷ�е�һ������ʱ���������ʵز�ίԱ�������ϣ�Ҳ��ʯ̿��ǰ�ڡ��������ȼ͵������Լλ��359��318������ǰ��������������������ѧ�����һ������ѧ�ҿ��������Ҵ�����ʶ���������ȼͣ���Ȼȷ����ʼ���ս�ʱ����δȷ�����������ȼ��Ǹ����������������Ⱥ��������ģ���Ϊ�������ȼ͵��Ҳ�㷺�ֲ������ �������������ȼ��Ҳ�������ຣ��ʯ���ң�������ڱ�Ϧ�����Ǽ��������֮�䡣 ������� ���ʵز�ίԱ�Ὣ�������ȼͻ��ֳ�3���ף� ͼ���νף�Tournaisian�� ά�ܽף�Visean�� л���ջ���ף�Serpukhovian�� ŷ��ѧ��ͨ����ʯ̿�ͻ��ֳɵ��ϼ����������Ǽͣ���������ѧ�����������ȼͻ��ֳ�4���Σ� ��º��˽ף�ʱ���൱�ڽ�ͼ���ν�����֮����ʱ�ڣ� ŷ�����ף�ʱ���൱�ڽ�ͼ���ν�������ά�ܽף� ÷��÷�˽ף�ʱ���൱��ά�ܽ����ڣ� ��˹�ؽף�ά�ܽ�������л���ջ���ף� ��Ϧ����������Ϧ����������Ӣ�Pennsylvanian���ǵ�����ʷ�е�һ������ʱ���������ʵز�ίԱ�������ϣ�Ҳ��ʯ̿�ͺ��ڡ���Ϧ�����Ǽ͵������Լλ��318.1�� 1.3��299�� 0.8������ǰ��������������������ѧ�����һ������ѧ�ҿ��������Ҵ�����ʶ����Ϧ�����Ǽͣ���Ȼȷ����ʼ���ս�ʱ����δȷ������Ϧ�����Ǽ��Ǹ��������ı�Ϧ���������������ģ���Ϊ��Ϧ�����Ǽ͵��Ҳ�㷺�ֲ������ �����ִ�����Ƶ����ﶼ�����ڱ�Ϧ�����Ǽ͵��С� ������� ��Ϧ�����Ǽ�ӵ�����ͬ��������ַ�ʽ�����ʵز�ίԱ����ݶ���˹��ѧ��ķ�ʽ���仮�ֳ�4���ף� ��ʲ�����ף�Bashkirian��323.2-315.2������ǰ Ī˹�ƽף�Moscovian��315.2-307.0������ǰ ����Ī��ף�Kasimovian��307.0-303.7������ǰ ���ȶ��ף�Gzhelian��303.7-298.9������ǰ ŷ��ѧ��ͨ����ʯ̿�ͻ��ֳɵ��ϼ����������Ǽͣ������������Ǽ���ʼʱ��ȱ�Ϧ�����Ǽ��磬�����ֳ�3���ף� ���Ѷ��ף�ʱ���൱��л���ջ��������ʲ���������ڣ� �������Žף�ʱ���൱�ڰ�ʲ����������������Ī��ף� ʷ�ٷҽף�ʱ���൱�ڸ��ȶ��ף� ������������ 299-251������ǰ ȫʱ��ƽ������O2���� Լ23 Vol %��Ϊ�ִ���115% �� ȫʱ��ƽ������CO2���� Լ900 ppm��Ϊǰ��ҵʱ��3���� ȫʱ��ƽ���ر��¶� Լ16�棨�����ִ�2�棩 ��ƽ�棨�����ִ��� �ڶ�������������ȶ�����60�ף�200Ӣ�ߣ����������½���ĩ���ȶ���-20�ף�-66Ӣ�ߣ� �����ͣ�Permian����Լ2.9-2.5����ǰ�����������һ������ʱ������ʯ̿�͡�������֮�䡣��������͵���ʯ���DZȽϷ����ģ�����ʼ�������ľ�ȷ���ȴ�����顣�䲻��ȷ�ȿɴ��������ꡣ������������Ϊ���֣�Ŀǰ������ʹ�����ַ���Cisuralian��Guadalupian����ƽ����Lopingian����PermianԴ�Զ���˹�ı˶�ķ�ݡ�������Ϊ������һ˵���ڵ¹���ͬ����ز���ϲ���þ�ʻ��ң��²��Ǻ�ɫɰ��֮�ʡ� ���� ���ڵ�ֲ������ާ������ާΪ���������нϴ�仯����ľ�ࡢ«ľ�ࡢ����ާ���´���������˥���������֮�Խ��ͺ�������ֲ��ɰ����Ϊ���ӣ������ʼ��չ�� �����������ʢ����������ռ���ƣ����嶯��Ҳ����Ҫ���֣���ʯ�������Է��죻̦����˥�ˣ���Ҷ��������������濪ʼѸ�ٷ�չ���Ӵ����ƬĿ���ܶ����һ����չ����Ȼռ��ͳ�ε�λ�����ж����״δ�����ʢ������Ŀ��ʼ��Ŀ�������ڣ�����Ŀ����Ŀ�ȺϹ��ٶ���������ơ� ������ĩ�����˶�����-����������¼���90%��95%�ĺ����������������ϸԭ��Ŀǰ�в���ȷ�� ���� ���������ں��䡢�����㲼�����ں��ˡ��ɺ��������������ϲ���ӡ�Ȱ뵺���ͻ�˹̹���롢�Ĵ����ǡ��ϼ��ޡ��й������ϲ��ȵ��б����ҡ���ˮ������ ���ڻ�ɽ��㷺���������ڳ��š������İ����������˶����Ƕ�����ĩǿ�ҵĹ����˶��������ĿƵ�������ײ���������ؿ��˶��У�����ǿ�ҵĻ�ɽ�������������ز���������������¡��ŷ��½����Ϊһ�塣���Ǻ��й����������ϲ��İ����ײ���ڶ����;�����һ�����塢���ʡ���ɽ����������������뼰�С��������ҡ������ҵ������ ����˹���������ڱ�γ30��-40�㣬�Ե��к�������ӡ�������ǣ���֧�ذĴ���������������γ30�㣻����֧�븲���й���½�������������ձ���ɢС�����ͨ���������������ຣ��ͨ��������ĩ������ĺ��ˣ�ʹ����ֺ����������������й����ϡ��ͻ�˹̹������һ�������������ͼ䱣�ֺ�����  |
| ������ͣ� | |
| W(��15����Ϣ) |
|
| �����Ϣ�� | |
| �������� ������ϵ������ Э������ʱ ǰ����� ���ǹ��� | ���嶯�� ������ �Ǽ����� ��С���Ǵ� ������ |