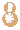| ��ҳ ==> �ܵĹ��� ==> �鿴��Ϣ |
 �������� ���������� ��������� Eukaryota �磺 ����� Animalia �ǽ磺 ���������ǽ� Parazoa Grant & Todd, 1838 �� ������� Porifera ���������Ƕ�������һ�����ϵ��ǽ磬�������ë������Ȳ�ͬ�������к�۵Ľṹ���ֻ���ϸ�������ֲ������������һ������֯������Ψһ�����ŵIJ�������ֻ�к��࣬���ڶ�����š���ЩҲ����������̶����ţ�����ֻ��˿�̳�ͬһ�����֡����������Dz��ԳƵģ����������еĶ��������ǻ���һЩ�ĶԳơ������������ڰ�������ǧ������֣�Լ��150�������ڵ�ˮ� ��������磺 ����� Animalia �ǽ磺 ���������ǽ� Parazoa �ţ� ������� Porifera Grant in Todd, 1836 �� ���ʺ���� Calcarea ���ź���� Hexactinellida Ѱ������� Demospongiae ��׳�� Stromatoporata ������ţ�ѧ����Porifera��Ϊԭʼ�Ķ�ϸ�����Ҳ�ƺ��ද���ţ�Spongiatia��Spongia����һ���Ϊ���ࡣ����û����������ѭ��ϵͳ���෴�����Ǵ��������ˮ�����Լ������������ʳ���������������������е���ˮ���� ������״��ֲ����Ҹ�ֲ�ں����У����Ժ���ǧ������������Ϊ��ֲ��������������Ǽ����߱�����������Ķ������������Ե�ʮ���������ڲű����붯�����ͷ�������ڻ��ڻ�������Ѷ��ϵͳ����ѧ�ĸ���֤�ݣ�����Ӧ������DZ����ڶ���绹�Ƕ��������Գ�һ�������۵�������ѧ�����ڲ��������С� ���ද��������кܶ�С�ף���ˮ�ף��������һ���д���ڡ� ��������Ͷ��ֶ�����С�IJ������ˣ����ȴ��45������ĺ���������4.2�ס��������ɫͬ���Ƿḻ��ʡ� ϸ�����ѿ�ʼ�ֻ�����δ�γ���֯�����٣�Ҳû���γ��������߲㣨�����߲㡢���߲�����߲㣩�� ������ڡ�������ϸ�����ɡ����ϸ����ƽ����Ƥ�㣨��ϸ���㣩���ڲ�ϸ����θ�㣨��ϸ���㣩�����б�ë����������ԭ�����죬�ơ���ϸ������choanocytes������Ҫ����ʳ��ϸ�������������ã���ˮ��ͨ�����ڵĹ���������ϸ����ɱ�ë�Һͳ�ˮ����ɸ��ӵĹ���ϵͳ������ʳ���ĺ�ˮ�������ڲ�ϸ����ë�IJ���������ˮ���������ڣ��������Ķ������ź�ˮ�Ӷ��˵ij�ˮ���ų����⡣ ���ڡ�������ϸ��֮�䣬����һ���н��㣬����������γ������ϸ������ֳϸ�������ϸ��������˿ϸ���ȡ� ���ද������ڶ����֧�ֵ���״���������ڹ�����Endoskeleton�����ɽ�ԭ���ף�collagen����̼��ƣ�CaCO3���Ͷ������裨SiO2����ɡ� ����û����ϵͳ��������ϸ����ͬ��ʳ���ֹ����������Ա���Ϊ�Ƕ���������γɵĿ�ʼ�� ���Ϊ����ͬ�壨Hermaphroditic) ���� ���ʺ���� ��Calcispongea�� ���ź���� ��Hexactinellida�� Ѱ������� ��Demospongiae�� ��׳�� ��Stromatoporata�� �ɷ����У�������ſɷ�Ϊ����һЩ�٣� ��ͨ����� ��Demospongea�� ��������� ��Hyalospongea�� ���ʺ���� ��Calcispongea�� Ӳ����� ��Sclerospongea�� ��������������������һ�ֶ��⺣������������ֽ���Ȼ������������˻��˵ı��ζ�����Ƕ�����չ���ġ�һ����ԣ���ЩϸС������������������ɰ�����һ��������ֳϸ���ϵ���ë��Ƥϸ������ɵġ���ʮ��ǰ���������ﱻ����Ϊһ���ţ���������ϵ����ѧ���о�ȴ��ʾ��������������������ʵ�Ƕ�ϵȺ�ģ��༴������������������ص���Ⱥ����ɵġ� �����������Ÿ��ӵ��������ڣ��������Ժ����ԵĽμ佻�档��ͷ���ද����������������Ƴ�һ�㣬�ɹ̶���������ϸ����ɵ���������壩����������ĺ���������ϸ������Щϸ��������µ������塣֮����������ת��������壬�����Է�չ���Է�չ��ʼ�����������ϸ����һ������ infusorigen �������ڣ��ڴ˴���չ�����װ�״�ľ���������ϸ����ת����ѡ���������ϸ���г�ȥ�������ѽ�϶��γɵ��׳棬Ȼ���׳������������������ٴα������״�Ķ����Ⱦ�� ��������һ�ȱ���Ϊ��ԭ������Ͷ���֮���ݻ���һ��������̬�������ڣ���������Ϊ���˻��˻���˵Ķ�����ǵ���ë�׳���������ë�׳����ƣ������ǵ��ڲ���ֳҲ�����������ڷ������������ơ��������� �ġ�DNA �нϵ͵�GC������40%������һ��ֵ����ë�����ƣ�����ë��ͨ����˫ϸ���ˡ�����������������صĻ��������ڶ��Ŧ�ζ�������г����Ⱥ�� ����������������Ҫ����Ⱥ�����ζ����ź�ֱӾ�����ţ���ʱҲ�������̶����ź͵��߶����š� ���߶�����ֻ����һ��ֻ��ʮ������ʱ�й�һ�����������֣����Դ˱㲻���ټ�������ˣ������˻���������һ����ʵ����Ⱥ���������������ֶ���ֻ�е�һ�����֯�� ���ζ����� ���ζ����ŷ�����ͷ��ٵ�ԭ�����ڡ������Զ�ʮ����ʮ��ϸ���ų������ij��ȣ���Щϸ������ǰ�˵ı�Ե������һ������״��֮Ϊ��ϸ������ֳϸ������ϸ���������Եط�չ�ɳ�״�����壬Ҳ���������Ѻ;��ӣ��������ܾ���������ë���׳档 ���ζ������������������߳����� Pseudicyema �� Dicyemennea �� ֱӾ������ ֱӾ�����ŷ��������ຣ��������������϶�ڣ�����֯��϶����ֳ�١����ҵȡ����ಡԭ��ᵼ�²�ͬ���ֵ�����ɥʧ��ֳ������ ��֪����ֱӾ���Ŷ���Ϊ��β�ļ����档 ����������磺 ����� Animalia �ǽ磺 ����������ǽ� Eumetazoa Butschli, 1910 ��֧ ����Գƶ��� Radiata ����Գƶ��� Bilateria ��������ѧ����Eumetazoa����ָ���о���ϸ����֯�Ķ��������ֻ�в�����������������е������Ų������������� ���������ϸ���������ӣ�������ν��"��������"�����ǵ���̥���ٻᷢ�������߲㣺���߲�����߲㡣ֻ�����������ž����ػ���ϸ����������ϸ���� �����������ٷ�Ϊ����Գƶ���ͷ���Գƶ�����߰����̰����� (Cnidaria)����ˮĸ������ (Ctenophora)�������ַ��ಢ���ܷ�ӳ��ʵ����ϵ������������Ե��֧���෨����ˮĸ���������Գƶ����и������Ե��ϵ�� ����Գƶ����磺 ����� Animalia �ǽ磺 ����������ǽ� Eumetazoa �ݻ�֧�� ����Գƶ��� Bilateria �ݻ�֧ ��ڶ��� Deuterostomia ԭ�ڶ��� Protostomia ����Գƶ��ѧ��Bilateria�����������ҶԳƶ�������Գƶ��Radiata����ͬ�����������Eumetazoa���� ����Գƶ���������������߲㷢����������ӵ�����߲㡣���߲��������߲��ݻ������ġ�������Գƶ���ȷ���Գƶ������ ����˼�������������dz�����Գơ�������ǰ�ο�ʼ�γ��Բ���Cephalisation����ijЩ�������״���⣨�����ᵼ�¸ô�������խ�����⣬���м����������¸ô����ɸ�ԭ�����ڡ����Ǵ־��г��ij����ѪҺϵͳ���ڱ�Ƥ��Epidermis���ͳ�֮������ǻ��Coelom���������ڲ��������Һ���γɾ�ˮѹ��������̬����������ѪҺϵͳ���������й��ͨ��ԭ���ͺ�����ɡ� �������������ѷ�ʽ��������ԭ�ڶ���Protostomia�������䣨��ڶ���Deuterostomia��������Գƣ�ԭ�ڶ����ڶ���� ��ڶ����磺 ����� Animalia �ǽ磺 ����������ǽ� Eumetazoa �ݻ�֧�� ����Գƶ��� Bilateria ���ţ� ��ڶ������� Deuterostomia Grobben, 1908 �� ���������� Chordata ���������� Hemichordata ��Ƥ������ Echinodermata ���ж����� Xenoturbellida ��ǻ������ Acoelomorpha �ų涯���� Vetulicolia ��ڶ������ţ�������ѧ����Deuterostomia���Ƕ����е�һ�����ţ�������������ǽ������Գƶ����֧�У���ԭ�ڶ�����ԡ���ڶ����ԭ�ڶ����������̥���������֣��ں�ڶ����У���һ�����ڣ��߿ף�����˸��ţ�����ԭ�ڶ�������������͡� ��ڶ��ﹲ������ִ���ţ� ���������� Chordata �����������ͬ�ࣩ ��Ƥ�����š�Echinodermata �����ǡ������ͺ��εȣ� ���������š�Hemichordata ����������ʯ����Ҳ�ǣ� ���ж����š�Xenoturbellida ����������Ƴ�����֣� ��ǻ�����š�Acoelomorpha ë���ſ���Ҳ���ں�ڶ�����������Ⱥ�������ų涯���š� ��ڶ����ǰ�ڶ�����ܾ���һ��ʼ���ᷢ����һ����ϸ����ɵ�С��֮Ϊ���ߡ��ں�ڶ����У����ڵIJ��ֻ�ƽ�л�ֱ���ᣬ���Ϊ�����ѸҲ�������ijЩԭ�ڶ����ϣ��紥�ֹڶ��������ĺ�ڶ�����в����Ѹ��ϸ������̥�еķ�չ������ĸϸ���������ġ���ˣ�����һ��ʼ���ĸ�ϸ���ֿ���ÿһ��ϸ�����ܹ��γ�һ�����������壬����һ��ϸ�����Ƴ������ߣ�������ϸ���ͻᲹ������� ��������ͼ������ﶼ�����ѣ���ԭʼ��ʯ�еļ�Ƥ����Ҳ��ʾ�������ѵļǺš������м����������β�����������ĽΣ�֮�У��������пյ�������һЩ�İ�������Ҳ���й�״������������̥����ʱ���������Ǽ���������п���������Ϊ��Ƥ�������˻�����ϵͳ���ֱ��������Ƿ�Ҳ������Dz����ܵģ�����������ͬ����ʵ���������൱�Ŀ����ԣ������ִ�ĺ�ڶ��ﶼ�ݻ���һ�������ѡ��п������ͷֶε����塣��һ����Ӧ���ƺ����ʱ�������ų涯���С��ڶ�����Ⱥ�� ԭ�ڶ����磺 ����� Animalia �ǽ磺 ����������ǽ� Eumetazoa �ݻ�֧�� ����Գƶ��� Bilateria ��δ�ּ��� ԭ�ڶ��� Protostomia ���� ��Ƥ�������� Ecdysozoa ���ֶ������� Lophotrochozoa ԭ�ڶ��������ѧ����Protostomia��������������������̥ʱ�ڵ�ԭ�ڻᷢչΪ����Ŀڣ����������������γɵġ���Ƥ���Ecdysozoa�����ֶ��Lophotrochozoa��������ԭ�ڶ�������ѷ�ʽΪ����ʽ�����䲻���ȷ��ѻᵼ���м���һ��ʱ�ڣ������ĸ���ϸ����Macromere�����ĸ�Сϸ����Micromere�������ද����������ǻ������ǻ�������߲��γɵġ� �������ڶ����������ο���ڶ�����Ŀ�� ԭ�ڶ�������� ��Ƥ���Ecdysozoa�� ���Ƕ����ţ�Kinorhyncha�� ��ҷ�����ţ�Priapulida�� �߳涯���ţ�Nematoda�� ���ζ����ţ�Nematomorpha�� ��צ�����ţ�Onychophora�� �������ţ�Loricifera�� ���������ţ�Tardigrada�� ��֫�����ţ�Arthropoda�� ���ֶ��Lophotrochozoa�� ���嶯���ţ�Mollusca�� ���ڶ����ţ�Annelida�� ̦�����ţ�Bryozoa�� ���㶯���ţ�Brachiopoda�� ��涯�Platyzoa�������������ţ���ֱ�ӹ��ൽ���ֶ�� ���ζ����ţ�Platyhelminthes�� ��ë�����ţ�Gastrotricha�� �θ�����ţ�Gnathostomulida�� ���ţ�Micrognathozoa�� ���ζ����ţ�Rotatoria�� ���ڶ����ţ�Cycliophora�� ��ͷ�����ţ�Acanthocephala�� ��Ƥ�����磺 ����� Animalia �ǽ磺 ����������ǽ� Eumetazoa ��δ�ּ��� ����Գƶ��� Bilateria ԭ�ڶ��� Protostomia ���ţ� ��Ƥ�������� Ecdysozoa Aguinaldo et al., 1997 �� ����֫����(Panarthropoda) ��֫������(Arthropoda) ��צ������(Onychophora) ����������(Tardigrada) Scalidophora ���Ƕ�����(Kinorhyncha) ��ҷ������(Priapulida) ��������(Loricifera) �߳涯����(Nematoda) ���ζ�����(Nematomorpha) ��Ƥ���Ecdysozoa����һ����ԭ�ڶ��������֫�����š��߳涯���źͼ���С�š������Auinaldo������1997�궨�壬��Ҫ������18S ������RNA�������������ͬʱҲ��һϵ����̬ѧ֤����֧�֣���Ϊ���������������ɵ�������Ķ������Ƥ(ecdysis)��������Ƥ����ӽ��Ķ���Ҳ������Perrier��1897���Seurat��1920���������̬������ ��Ƥ�����������Ĺ�ͬ������һ������ı�Ƥ��Ӣ��cuticle�������л�����ɣ��ܹ����Ŷ��������������ɵ�����Ƥ�����ɴ˵�������Ƥ����ȱ���˶���ë���������γ������ӣ�����̥��������ԭ�ڶ���һ���������ѡ���Ƥ�����һЩ��֧��������һЩ��ͬ���������绺��������߳涯�ﶼ��һ��������ԳƵ���(pharynx)�� ��Ƥ������������ţ���֫�����š���צ�����š����������š����Ƕ����š���ҷ�����š��������š��߳涯���ź����ζ����š�����һЩ���࣬���縹ë�����ţ�������Ϊ���������е�һԱ��������ȱ����Ƥ�������Ҫ����������ͨ��������������֫����Ĺ�ͬ�����Ǿ��зֽڵ����壬ԭ����Ϊ�ɻ��ڶ����ݱ������������һ����ɷֽڶ���(Articulata)��Ȼ�����Dz�������̫��Ĺ�ͬ���������ڿ������Ƿֱ��ݻ����˷ֽڵ��ص㡣���⣬��18S ������RNA����������ڶ����е�ë���ſ����������ڶ��ﲻͬ��������Ƥ�����Ƥ�����в�������֫����ij�Ա��������ɻ�����(Cycloneuralia)����ͨ������Ϊ�Dz�ϵȺ�� ���ֶ����磺 ����� Animalia �ǽ磺 ����������ǽ� Eumetazoa ��δ�ּ��� ����Գƶ��� Bilateria ԭ�ڶ��� Protostomia ���ţ� ���ֶ������� Lophotrochozoa ���ֶ��ѧ��Lophotrochozoa���Ƕ�����е�һ��֧����������Գƶ������Ƥ���Ecdysozoa�����ԭ�ڶ��ԭ�ڶ���ͺ�ڶ��Deuterostomia������Ϊ����Գƶ����������֧�������ɴ��ֹڶ��Lophophorata���͵��ֶ��Trochozoa���������϶��ɡ� ���ֶ��������ͳ�ϵģ� ���嶯���ţ�Mollusca�� ���ڶ����ţ�Annelida�� Ŧ�ζ����ţ�Nemertea�� �dz涯���ţ�Sipunculida�� Μ�涯���ţ�Echiura�� �������ţ�Pogonophora�� ���ֹڶ������̦�����ţ�Bryozoa�����ڸض����ţ�Entoprocta�������㶯���ţ�Brachiopoda������涯���ţ�Phoronida������ �������18S rRNA���У����ζ����ţ�Rotifera�����������������ζ�����½����ֵĻ��ڶ����ţ�Cycliophora��Ҳ�ܿ������ڹ��ֶ�� ���й��ֶ�������ԭ�ڶ��Protostomia����������̥�ڣ��ڵķ����ڸ��ŷ���֮ǰ�γɡ����ֶ��ᄀ����״�����ܶ࣬��ͨ��Ϊ���״������������ ���ֶ��� ��ͳ���������ൣ�ֶ��������嶯��ͻ��ڶ�����ֶ��������ʷ����һЩ�ؼ��Ĺ��ԡ��׳�������ΪС����Ӿ�壬��Ϊ�����׳档�����׳��������в�ӵ��������ë����������Ӿ�Ͳ���ʳ����ڡ����ˡ���һ�Գ��ı�ë�����ֶ�����׳�dz����ƣ��������������ܴ�ֱ�����������һֱ��Ϊ��֫����ͻ��ڶ����ǽ��ף���ԭ���������ද�ﶼ������ֽڵ�������֫���ﲻ���е����׳棬��������ѧҲû��֧����һ�۵��֤�ݡ� ���ֹڶ��� ���ֹڶ�����������Ϊ�ǵ�ϵȺ��Monophyly����������涯���ض�������㶯����к�����ӵ�д�����ʯ��¼�����Ƕ����д��ֹڣ�Lophophore���������пյĴ��֣����ڲ�ʳ������������Ϊ���ֹڶ���ͺ�ڶ��������й����������µ��о������뵣�ֶ���������һ�� �½����о����֣����۵��ֶ�����ֹڶ��ﶼ���ǵ�ϵȺ�������������嶯��ͻ��ڶ��������ŵĸ������ݻ����Ϸ�����ͬ��֧�����价�ڶ���Ķ�ë�٣�Polychaeta���ĸ�����֧���ڹ��ֶ���Ļ���������Ϊ��ϵȺ��Ӣ��paraphyly���������˹��ֶ����ԭʼ��̬������С�ſ���Ϊ�ػ�����Ⱥ�����������Ҫ��һ�����ۡ����ڹ��ֶ����ڲ���������ݻ������ж�����̬ѧ�����Լ���������ѧ�����������в���ȷ���� ��涯���磺 ����� Animalia ���ţ� ��涯������ Platyzoa ��涯��ԭ���DZ��ζ����ŵ�ͬ��ʣ��˴��Ǽ����ŵ�ͳ�ƣ���������Գƶ����涯������(Platyzoa)�����������࣬���˷�����δ��Ϊ��ʶ�� ���ζ����� (Platyhelminthes) ��ë������ (Gastrotricha) �θ������ (Gnathostomulida) ���� (Micrognathozoa) ���ζ����� (Rotatoria) ��ͷ������ (Acanthocephala) ���ڶ����� (Cycliophora)�����ܣ� ���ζ����ë����û����ǻ��������������м���ǻ�����ǵ����ʹ������ơ������м�������ļ�ͷ����û����Щ�ṹ�������γ�һ����ϵȺ����Gnathifera�� ��涯�����ŵĶ���͵��ֶ����ǽ��ף���ʱ���鵽���ߡ���������������Ѷ���(Spiralia)������ı��ζ�����ͨ��Ϊ����ƽ����棬������������ĺͼ��������ࡣ���ں�ˮ����ˮ��½�ض����������㲼��������ء��峤��1�������ɷ�֮һ��15�ס���3�����ͣ��г桢������г档�����������Գƣ����ֽڣ�������ƽ������ϵͳ��������ѭ��ϵͳ����ǻ������֮��Ŀ�϶Ϊ����Ľ����֯��䡣 ����Գƶ����磺 ����� Animalia �ǽ磺 ��������� Eumetazoa �� �̰������� Cnidaria ��ˮĸ������ Ctenophora ����Գƶ��ѧ��Radiata��������Գƶ���(Bilateria)��ͬ������������(Eumetazoa)�� ��Щ���������Ϊ���λ�Ͱ�Ρ� ����Գƶ��������������߲㹹�ɡ��м���һ�н���(Mesogloea)�� �̰��������磺 ����� Animalia ��δ�ּ��� ����Գƶ��� Radiata �ţ� ��˿�������� Cnidaria Hatschek, 1888 �� ɺ���� Anthozoa ��ˮĸ�� Scyphozoa ʮ��ˮĸ�� Staurozoa ����ˮĸ�� Cubozoa ���ڼ���ˮ��� Polypodiozoa ˮ�� Hydrozoa ��ˮĸ�� Dipleurozoa ԭˮĸ�� Protomedusae ˮ�ʯ��(��) Hydroconozoa �̰������ţ�Cnidaria ����������˿�������ţ��Ƕ�����һ���š�������������Ϊ��ˮ�����⣬��������־�Ϊ��������������dz������Щ������ִ������Լ��11000�֡��̰�������������ˮĸ����һ�����ǻ�������ţ�����ˮĸ���������һ�š� ��˿�������Ŷ����������ص㣺 ��Ϊ��ʳ�ԣ��ٲ����������������������ڹ��������й�����õ����ġ� �����ɳʷ���Գƣ�ˮ�������������� ���������ϸ����ɣ���Ƥ�ͳ���Ƥ������֮����һ������״���н��㣬��֧�����á� �г�ǻ����һΧ�ڲ���Peristom�������ǿ�Ҳ�Ǹ��š� �г���20�ֵĴ̰�����˿���к��д�˿�ҡ���˿��һ�˵�����ͻ��Ĵ��룬�ܴ̼�ʱ�������˿���ſա���˿���ı�����ͻ�����������й�״�ṹ���̰����и߶�����������ʣ���ѹ�������»��ͷš� �ڳ�ǻ�н��а������� ��ɢ����ϵͳ������״ ˮĸ���ио����٣��ܸ��ܹ������ �д���ͬ������� ˮĸ��ɺ����������ˮ������Ϊ��˿����� ��ˮĸ�磺 ����� Animalia �ţ� ��ˮĸ������ Ctenophora Eschscholtz��1829 �� ���ָ� Tentaculata ���ָ� Nuda Ӳ�Ǹ� Scleroctenophora ��ˮĸ��ѧ����Ctenophora�������������ң���һ�����߲㶯�������Գƶ���ֱ�����Ϊ��ˮĸ�����š�ԭ�ʹ̰�����һ�����ǻ�������ţ���Ϊ�̰����ţ��ֳַ��� �нϷ�����н��㣬ӵ�м���״��ϵͳ�ͼ���ϵͳ�� ���߲㣻ɡԵ���ӣ���������������������Ρ���Բ�Ρ���ƽ�εȣ���������ΰ����г����е���ë��8��������û�д�ϸ�����������ҶԳƻ�ʷ���Գƣ����ʺ��������Ӿʱ���ΰ壻���ڼ���һƽ��о�����û��θ˿�� ����ѧ�ϣ���ˮĸ��ˮĸΪ��ͬ�ţ�����ˮĸ���̰���ͨ��ȫ�������������ˮĸ������ϵĶ�ϸ��������ܶ�ṹԶ����ˮĸ������纣�������ţ�Ҳ�ж�����ţ��Լ����������Щ���ﶼ��һ����ϸ���ֻ���û����֯�ֻ���û���γ��߲㣬û����ϵͳ��û�м���ϵͳ����������ߵ������ϵ�������ӻ������ת¼�����������Ķ�ϸ����������û����ϵͳ�ͼ���ϵͳ����ˮĸ��ˮĸ��������ͬ���������߶����Ľ���������ϵͳ�� ��ˮĸ�������������Ļ������Ʋ�ͬ��ˮĸ�����磬ˮĸ�������ߵȶ�������ϵͳ�����ؼ���������ˮĸ�ж������ڡ�ͬ������ˮĸҲ��ʹ��ˮĸ���������ﳣ�õ����ʣ����Ͱ����������Ѫ���صȡ��ڴ�ͬʱ����ˮĸ����ʹ�ùȰ�����Ϊ����Ҫ�����ʣ�������������ͬ��ʹ�ùȰ�����Ϊ���ʣ�����ˮĸ�йȰ����������Զ����������Ķࡣ���������������ϵͳ������ԭʼˮĸ����ϵͳ���������ģ�ӵ����ˮĸ��ͬ�Ļ������ƣ���˿�����Ϊ��ˮĸ����ϵͳ�Ƕ��������������ȫ�µ���ϵͳ�� ��ˮĸ�ſ��Է�Ϊ3����5��Ŀ�� ���ָ٣�Tentaculata�����д��ֵ�ˮĸ�� ����ˮĸĿ��Cydippida�����������ˮĸ��Pleurobrachia���� ��ˮĸĿ��Lobata��������Mnemiopsis�� ����ˮĸĿ��Cestida�����������ˮĸ��Cestum���� ����ˮĸĿ��Platyctenea�����������ˮĸ��Ctenoplana���� ���ָ٣�Nuda�������ֵ�ˮĸ�� ��ˮĸĿ��Beroda���������ˮĸ��Beroe���� Ӳ�Ǹ٣�Scleroctenophora�� ���嶯���� ����� Eukarya �磺 ����� Animalia �ǽ磺 ����������ǽ� Eumetazoa ��δ�ּ��� ����Գƶ��� Bilateria ԭ�ڶ��� Protostomia ���ţ� ���ֶ������� Lophotrochozoa �ţ� ���嶯���� Mollusca ���� Polyplacophora ����� Monoplacophora ˫�Ǹ� Bivalvia ����� Scaphopoda ����� Gastropoda ͷ��� Cephalopoda Ǹ� Rostroconchia ̫��Ů���ݸ� Helcionelloida ���ʯ�� Tentaculita �����ݸ� Hyolitha ���嶯���ţ�ѧ����Mollusca������������������ֶ����Զ��ԣ��Ƕ����ĵڶ����ţ������ڽ�֫�����ţ�����ʮ����֡����嶯������Ӧ���ͬ�������ֲ��㷺���Ӻ������´����ȴ����Ӻ��Ӵ�����������ƽԭ����ɽ��½�ء���ˮ����ˮ�ж��д�����Ա��������ţ���Ӱ������ݡ����������֡� ���嶯����̬��ϰ�Բ��������������嶯�������������չ���ɴ�12�ף���С������ȴ����1���׳������������й�ͬ�Ļ����������������ڹ�����������������ֽڣ�����ṹ�ɷ�Ϊͷ���㡢�����ź�����Ĥ4�����֡��������嶯�������Ĥ����ڳ����ʵ�Ӳ�DZ������塣����ģ����״���������ͬ�����˳����ڵĸ��㶯��֮�⣬���嶯��ĵĿ��嶼�����ҶԳƵġ� ���嶯�����пǣ������ݡ��ĸ�ȱ��ࣻ������½���ϵ�������ţ�����������㡢���������������������ʧ�����嶯�������һ�������ǰ�������Դ��ƶ��Լ������壬�ܶ��һ�����Ƶ����������������������������塣 ���嶯������Ҫ�������������������������ڹ������ֳ����ҶԳơ����ֽڣ��������Ƥ�����Ա��������γ�����Ĥ��Χȫ�������ܹ����ڱ����õ�ʯ���ʽ�ǣ�Ȼ���������嶯���������˻�(������������)���Dz������ڣ����������������ࡢ������ĿDZ����������ߣ����Կ��������С�����嶯��ĸ����м��������Ҳ�е������Ѿ��˻������˶����١� ��������嶯���ͷ�ϳ��к��۾�һ���Ĵ��ǣ�������֪��Χ������������м�ǧ��С�����ݣ���Ϊ���࣬�����Ƕ������е����٣��ɶ��н��ʳݰ���ɣ���״����ﱵ�����Ҫ������ʳ�����������������ë�����Լ���ˮ����˫������ô˹���ˮ�е�ʳ������� ˮ�������嶯����й����Ϊ�������ų��������أ����Ǻ�������Ϊ����������Ĥ�γ�λ������������֮�䣬���Դ���������ǻ��ˮ����ȡ������������ţ��½�������嶯����й����Ҳ�����������÷κ���������й���ᡣ ��Щ���嶯���Ǵ������壻��Щ�Ǵ���ͬ�塣ͷ��ټ����ָ���������ܾ�������ͬ�����������ܾ�����ͬ�����嶯���в�ͬ�IJ��ѷ�ʽ�������ࡢʯ̋�Ȱ���ֱ�Ӳ��ں�ˮ�У����ޱ�����ʩ����˲����������࣬����˳��������ȴ���ࡣ��Щ����������������������ڽ��ʰ�ס������������Ѵ��н��ʰ�ס����пǡ� ���ֵ����嶯����о��ü�ֵ����������ܲ����顣���������ֵ�ˮ���İ��࣬���ڵ������ʿ�����ť�ۡ����㡢���ۡ�ĵ�õȿɹ�����ʳ�ã������������嶯���ṩ���������ʳ����Դ�����嶯����˶��������棬�������ֶ������к������磺��������䴬�棬���ֱ��������ں�����ƻ���ֻ��������ľ������в��ֵ�ˮ�����嶯��Ǹ��Ρ��ձ�ס������м�������ᴫȾ������ ���嶯���������Ҫ�ɷ�Ϊͷ���㡢������3�����֣��������ﻹ���д�����Ĥ�γɵĿǡ� ͷ ͷλ�������ǰ�ˡ� ���� �����λ��ͷ������帹�棬�������������ɶ༡������ɵ��˶����١� ������ ������λ�����屳�棬����������ڰ�Χ���������١� ����Ĥ������ǻ ����Ĥ�����嶯�ﱳ����������������չ�γɵģ������������������ţ�������������Ĥ֮��Ŀ�ǻ��Ϊ����ǻ������ǻ���������ٵĿ��ڣ�����š����ס����ס���ֳ�ȡ�˫�Ǹ����������������嶼�����������ǻ�ڡ� �� ��Щ���嶯�������Ĥ�����̼��ơ������ʺͱ���Ӳ�����γ���ǣ���ǵ�������ǽ��ʲ㣬�в�������״����ʯ�����ڲ����Dz�״����ʯ���ֵ����嶯�����Ҫ����ʯ��ɣ��������������ĵ���ȴ�ɷ���ʯ��ɡ� ��ϵͳ ����˫�����������⣬�����嶯����������Ҫ�ĸ�������һ����Ҫ���ᵽ�����ţ���һ�����ᵽ���㡣������������Ҫ���ڶ��ڳ����²ࡣ �������嶯���д��ԣ����һ���ʳ������������嶯������۾�����ͷ������һ�Ծߴ��й��ܵĴ��ǣ��ɼ�⻯ѧ���ʡ����������ȡ� ����ϵͳ ����ϵͳ ��������嶯��ֻ��һ�������е�����ֻ��һ�����ֵ����γ���ë״����Щ���˿״�����嶯�������ʹˮ�ӵײ����붥���ų������ĵ�˿��������ë�䵥˿��������ë������һ����������Ĥ������ˮ��������������˿�ɱ���������ࡣ�������и������������(osphradia)���������к����ʻ�����ɳ��������Ĥʱ��������ë�仯ֹͣ������ֱ���к���������ʧ��ÿ�������������ᵽ�彫��Ѫ�ܣ�������ѪҺ�������Ѫ�ܡ� ѭ��ϵͳ �ֵ����嶯���ǿ���ʽѭ��ϵͳ����Ȼ���嶯�������߲㶯��������ǵ���ǻȴ�൱С����������������ֳ�١���ǻ�ڴ����Ѫǻ������ѪҺ����Һѭ�����������١���������ѪҺ����Ѫ�����ף����������ʱ�����Cu2+��������ǵ�ѪҺ������ɫ�����ǵ�������Щֻ��һ���ķ�����Щ�ж���ķ������Ǵ����õ�����Ѫ�����͵����ң�Ȼ�����͵��൱�̵�������������ȫ���� ���嶯����ķ����в��ֵ���й���ܡ��ķ�����ѪҺ�еķ�����ŵ���ǻ���γ���Һ��λ�ڱ�����һ�����ܣ��ɴ���Һ����ȡ���õ����ʡ��ų����õķ�����ɹܵ��ŵ����⡣ ����ϵͳ ��ֳϵͳ ���� ����ѧ�Ҹ������嶯���Ӳ�Ǻ�����ṹ�IJ��죬�����嶯��ֳ���10���٣��ֱ��ǵ���١����١��ް�١�����١�����١�˫�Ǹ١�Ǹ١�ͷ��١����ʯ�ٺ������ݸ١� ����� �ں��������ʱ�ѳ��֣����ұ�������������ǵ������ǿǵ���״��ñ�ӻ����ӡ� ���� ���������ں���������ѳ��֣����ǵ�����Գƣ�����Բ�Σ���8��ǰ���ɵı��ǣ���˳ƶ��١� �ް�� �ް�����ﲢû�йǰ�ǣ���˳�Ϊ�ް��������ǵ�������и�����״�Ľ�����Ƥ�����б������á����ǵ���������Գƣ�������Ƶġ�Ŀǰû���ҵ��ް������Ļ�ʯ���ִ����ﺣ����������� ������ �����������嶯��������һ�࣬��Լ��������ǧ���֣����ǵ����㶼�dz�������ĸ��棬��˱���Ϊ�����ࡣ��������������ţ�⣬���Ƕ����ݿǣ�����ֱ���Ϊ���ࡣ ����������ݿǣ���״����С����ɫ�������ͬ�����졣�����ݿ��ϵ����ƴ������˳ʱ�뷽��Ҳ���Դ�����Բ�ε��ݿ��ϣ��жϳ������С�����Ƕ��˵ĵط�����ԽϸԽ���ܵģ���;�Խ�� ½�س����ĸ����������ţ���������ݵȣ������еĸ�����϶࣬����������ҹ�ⱴ������ȶ��ǡ�Ϊ����Ӧ���ֻ��������ǵ��������������ط����У������ڳ��Һ���� ����ٴӺ�������ھͳ����ˣ�һֱ���ܵ��ִ���½�ء�����ˮ�ﶼ�����ǵķֲ��� ������ ����������˫������ֳ�˫�Ǹ٣��Ӱ������ȡ������Լ��ֵı��Ƕ�����˫�Ǹ١���������Ҫ���ص�����������ǰ꣬�����ǰ�֮���нºϹ��죻����ȱ�����Ե�ͷ���������ڸ����и�״��������˳�Ϊ�����ࡣ�����ں����ʱ�ͳ����ڵ����ϣ�����ˮ���ܷ������ǵĴ��ڡ� ͷ���� ͷ�������Ҫ���������Ǿ����߶ȼ��е�ͷ���������嶯���������������Ƕ��Ǻ����е���ʳ�Զ��ͷ��ٶ�������Գƣ�ͷ����������ǰ����ͷ�������з�����ۣ������н��ʵ��Ƭ���ڵ���Χ������һȦ�ܹ�������ʳ��������Ĵ��ֻ��������еľ�����ǡ��е������������ڲ����е�������ȫ�˻�������ʷ�ϣ�������ͷ�����������������ࡢ��ʯ����ʯ����ʯ�ȣ���������������������㡢�����ȡ�ͷ����ں����ʱ�Ѿ������ˡ� ������ ������븫��������Ե��ϵ�����ǵĽ��������״�ģ��ڷ���֮����Ȼ�Ƕ�ö������̬�����������гݱ��� �������������Գƣ�����Բ��״�����и���״��ǡ����Ǵ�������ں��ߵ���ɳ�У���Ҳ��������15,005Ӣ�ߵ���С����ǵ��㲿��ǰ�ˣ������������ǻ���ڡ�ͷ���������λ�������ͷ��ǰ�ˣ������㲿���档�����ܳ���U��״�����ſ���������ǻ��������һ�ԣ����ڴ��ڸ����Ժ�û��������������Ĥ����������Χ�д��ǣ��ио����á�����ϵͳ�������������ԡ��ࡢ�㡢��ȳɶ�֮�ᡣ���Ƕ��Ǵ������壬ֻ��һ����ֳ�٣���ֳϸ�����Ҳ������ų�֮�� Ǹ� Ǹ���һ���Ѿ����������٣������ں���͵����ռͣ����ǵĿǿ����������꣬�����ºϣ��γɡ���˫�ǡ���̬�ĵ��ǣ����������к������ؿǵȡ� ���ʯ�� ���ʯ����һ���Ѿ����������٣����ڰ��ռ͵�����ͣ�����ʷ���Գƣ����ǵ��εģ��������������ʯ�ȡ� �����ݸ� �����ݸ���һ���Ѿ����������٣���������ں���͵����ռͣ����ǵ����������Գƣ����е��εĿǣ����������������ݵȡ� ϰ�� ���嶯���ϰ����������죻��������½�ء���ˮ�ͺ�����зֲ���˫����ֻ�����ڵ�ˮ�ͺ����У�������Ⱥ�����������ں����С� �����ݶ������磺 ����� Animalia �ǽ磺 ���������ǽ� Metazoa ���ţ� ���ֶ������� Lophotrochozoa �ţ� ���嶯���� Mollusca �٣� �����ݶ����� Hyolitha �����ݶ����ţ�ѧ����Hyolitha���Ƕ�����һ���ţ���һ���Ѿ�����˵ĺ����пǵ���������仯ʯһ�㱣���ǣ��ڸǺ�֫�������֣����Ϊ���ʳɷ֣�����Գơ� ������һ�ද��ķ����ϣ�Ŀǰ��˵��硣�һ�������Ϊ�������嶯����֮�µ�һ���٣���һ���������Ϊ�����������ޱ����ϵ���ϵ��Ӧ��һ���������š� �����������ڴ��������ݶ����ţ��٣�����ֳ������٣�Ŀ���� ֱ���ݸ� �д������ݸ� ���ʯ�� ��������� Eukaryota �磺 ����� Animalia �ţ� ���嶯���� �٣� ���ʯ Ŀ�� ���ʯĿ��Tentaculitida�� �Ȼ���ʯĿ��Homoctenida�� ���߽�ʯĿ��Dacryoconarida�� ���ʯ��һ���Ѿ�����ĺ���������������δ��ȷ��Ŀǰͨ���������ʯ��ʯ�ĿDZڽṹ�ͳɷ��Լ�������̬�������ද����Ϊ���嶯�����е�һ�١� �������ʯ�DZڵĺ�ȡ�������̬�������ڽ����ʯ�ٻ���Ϊ3��Ŀ�� ���ʯĿ��Tentaculitida�� �Ȼ���ʯĿ��Homoctenida�� ���߽�ʯĿ��Dacryoconarida�� ̫��Ů���ݸ�̫��Ů���ݸ٣�Helcionelloida����һ��������Ĺ������嶯�̫��Ů���ݸ�����֪����ϵ��п��������Ҳ����˵����������һ������ǡ����²���������������Ϊ����١�̫��Ů���ݸ���John S Peel ��1991�꽨���� Ǹ��� ����� Eukarya �磺 ����� Animalia �ǽ磺 ����������ǽ� Eumetazoa ��δ�ּ��� ����Գƶ��� Bilateria ԭ�ڶ��� Protostomia ���ţ� ���ֶ������� Lophotrochozoa �ţ� ���嶯���� Mollusca �٣� Ǹ� Rostroconchia Ŀ �����Ŀ Ribeirioida ǿ׳��Ŀ Ischyrinioida ���Ŀ Conocardioida Ǹ٣�ѧ����Rostroconchia�������嶯�����е�һ���٣���һ�ٶ���Ŀǰĩ���������֣�ȫ��Ϊ��ʯ�֡����ٶ������̬�ӽ�˫���࣬�����ڷ���֮��������Ϊ��ͬ�����֡�����ʱ��֪���ǵĿ���������Ǵ�һ��ԭ���γ�һ��ԭ�ǣ��ٷ�����Ϊ����˫����̬�ij�����塣��ˣ�����һ����С���˫�ǡ���̬�ĵ������嶯�� ���ٶ���Ӻ�������ڳ��֣������������ھ���1972�걻��ʽ����ΪǸ١� Ŀǰȫ���緢�ֵ�Ǹٶ��ﻯʯ����40��������ۺ����ⲿ��̬���ڲ�������巢���������������Ϊ3Ŀ7�ƣ� �����Ŀ��Ribeirioida�� ����ǿƣ�Ribeiriidae�� ����ǿƣ�Technophoridae�� ǿ׳��Ŀ��Ischyrinioida�� ǿ׳�ǿƣ�Ischyriniidae�� ���Ŀ��Conocardioida�� ʼ���dz��ƣ�Eopteriacea�� ʼ���ǿƣ�Eopteriidae�� ��dz��ƣ�Conocardiacea�� ��ǿƣ�Conocardiidae�� ���Ͽǿƣ�Bransoniidae�� ���Ŀǿƣ�Hippocardiidae�� ������� ��������� Eukaryota �磺 ����� Animalia �ǽ磺 ����������ǽ� Eumetazoa ��δ�ּ��� ����Գƶ��� Bilateria ԭ�ڶ��� Protostomia ���ţ� ���ֶ������� Lophotrochozoa �ţ� ���嶯���� Mollusca Ŀ ����Ŀ��Tryblidioidea�� �ŵvĿ��Archinacelloidea�� ����Ŀ��Cyrtonellida�� ������Ŀ��Maikhanellidae�� Pelagiellida Tergomya ����٣�ѧ����Monoplacophora���������µ����٣������嶯���ŵ�һ����ϵȺ�٣���һ��ñ����״����ǣ�����ĵײ��������ٴ�����ǻ�ʯ���࣬��Ҫ���������������ز��С��ִ�7�֡�1952��������������״η��ָ��ද�����֣�������Ϊ��Ϊ���������µ�����������½����������������һЩ���֣����ǵ��ڲ������мٷֽ�������Ϊ����ѧ�߹������嶯���뻷�ڶ���ͬԴ��˵���� ��Ȼ�����ٵ��������ñ�������Σ����Dz����Ǹ����࣬����Ҳû�����й����� ���ٶ���ķ����бȽϴ����飬ͬһ���֣��ڲ�ͬ�IJο����ܻᱻ���벻ͬ�ķ��ࡣ����Ϊ�Ƚϳ����ķ��ࣺ ����Ŀ Tryblidioidea Pilsbry, in Zittel-Eastman, 1899 �����ܿ� Tryblidiidae von Zittel, 1899 ���ǿ� Cyrtonellidae Family Laevipilinidae Moskalev et al., 1983 ����� Monoplacophoridae Moskalev et al., 1983 �µ����� Neopilinidae Knight & Yochelson, 1958 ���ݿ� Tryblidiidae Pilsbry, in Zittel-Eastman, 1899 Family VemidaeMoskalev et al., 1983 ������Ŀ Scenellacea ���±��� Neomenioida խ��Ŀ Stenothecoidacea ��λδ�� Family Palaeacmaeidae ���ݲ�Ъ�غ���������ĸ��������(2005��)�������������λδ�����嶯���Ϊ��Щ����δ�ֱܷ浽���Ǹ���ٻ��ǵ���١� �����磺 ����� Animalia �ţ� ���嶯���� Mollusca ���ţ� ˫������ Aculifera �٣� ���� Polyplacophora ���٣�ѧ����Polyplacophora�������嶯���ŵ�һ���١��������������ں��У����дֵ����������ڳ�����ͳ��´�dzˮ������ʯ�ϣ���Ҳ��һЩ�����������ڽ����������Ϊ����Ȧһ�㶼�а�Ƭ��������֮Ӳ�ǣ���Ȧ���Ǽ�����֯�����ٰ�������ʯ��������˳���ij������ʯ�ϼ���������ʳ�á� ��ʱ�������ֵķ������������Sirenko (2006)�ķ��࣬�������£� ���� Polyplacophora de Blainville, 1816 ��ʯ���Ǹ� Paleoloricata Bergenhayn, 1955 Order Chelodida Bergenhayn, 1943 Order Septemchitonida Bergenhayn, 1955 ��ʯ���Ǹ� Neoloricata Shumacher, 1817 �۲�ʯ��Ŀ Lepidopleurida Thiele, 1910�������۲�ʯ�Lepidopleurus���� ʯ��Ŀ Chitonida Thiele, 1910�����������о�����������࣬���磺�����ʯ� ë��ʯ����Ŀ ����ʳ��λ���Բ�Σ���С��һ�����磺����ë��ʯ�� Ŀ��λδ����Incertae sedis�� Family Scanochitonidae Bergenhayn, 1955 Family Olingechitonidae Starobogatov et Sirenko, 1977 Family Haeggochitonidae Sirenko et Starobogatov, 1977 Family Ivoechitonidae Sirenko et Starobogatov, 1977 �ް���� ����� Eukarya �磺 ����� Animalia �ǽ磺 ����������ǽ� Eumetazoa ��δ�ּ��� ����Գƶ��� Bilateria ԭ�ڶ��� Protostomia ���ţ� ���ֶ������� Lophotrochozoa �ţ� ���嶯���� Mollusca �٣� �ް�� Aplacophora �ް�٣�ѧ����Aplacophora�������嶯���ŵ�һ���١�����250�����ҡ���һ����������״�����嶯� �������������ѧ���\����̬ѧΪ������֮ʱ���ް������������Ϊ������ͬ�࣬��һ�����1987��֮ǰ�ķ����п�������1987���𣬱�����õ�ȷ��Ϊ���嶯��֮һ�������Ϊһ�������ĸ١���ʱ����������Ⱥ��ɣ��ֱ�Ϊ�� �����Ǹ٣�Solenogastres�����������±�Ŀ��WoRMS�ѱ����������ټ��� βǻ�Ǹ٣�Caudofoveata��������ëƤ��Ŀ��ͬ����WoRMS��ѱ����������ټ� ������Ⱥ�����Ϊֻ�Dz�ϵȺ��������ķ�������֧������Ⱥ�ǵ�ϵȺ����˵�� �ӷ������ݼ���ʯ�ķ������ް�ٺܴ�����ǴӶ����ݻ���ֻ�Ǻ�����Щ���塱���˻��ˡ� ������磺 ����� Animalia �ţ� ���嶯���� Mollusca �٣� ����� Gastropoda Cuvier, 1797 �Ǹ٣� ֱ�����Ǹ١� �����Ǹ� ����٣�ѧ����Gastropoda�������嶯��������Ҫ����ɲ��֣�����ͨ����˵����ţ�����ݺ������������嶯���������ĸ٣�Լ60,000��80,000�����֡�����Լ409�������Ŀƣ������Լ202����ʯ�ơ��Ƕ����н����������(Insecta)�ĵڶ���١� ͷ����������зʺ���������㣬���Ե���������������IJ���Ťת��������Ҳ��Գƣ������н��һö���ǡ� �����ں�ˮ�еı���Ϊ���ݻ������������ڵ�ˮ�еı���Ϊ���ݻ����ϣ��Լ�������½���ϵ���ţ�������� ��������ĸ���ٵ����ֶ�����һ���������εġ��ǡ���������Σ�յ�ʱ��Ὣ�����������������С�Ҳ��һЩ���˻���ʧ��������ڲ�����ͨ������Ϊ������ �������ȫ���緶Χ�ڷֲ�����������Լ30,000�֣�����ˮ��Լ5,000�֣���½�أ�Լ30,000�֣��ȣ��ӿ����ϼ��ͱ����ĺ���������ȴ��������зֲ��� ������Ӧ�dz����������滷�����ӻ���ɭ�֡�ɳĮ��ɽ�����ӹ������������������Ӻӿڡ���̲��������ʯ�ij������ɳ�ij��´����������ЩӪ������ ˫�Ǹ��磺 ����� Animalia �ţ� ���嶯���� Mollusca �٣� ˫�Ǹ� Bivalvia Linnaeus, 1758 �Ǹ� ���г��Ǹ� (Palaeotaxodonta) �����Ǹ� (Cryptodonta) �����Ǹ� (Pteriomorphia) ������Ǹ� (Palaeoheterodonta) ����Ǹ� (Heterodonta) ���ʹ��Ǹ� (Anomalodesmata) ˫�Ǹ٣�ѧ����Bivalvia�������������ࣨLamellibranchia�������ࣨPelecypoda���������嶯���ŵ�һ���٣����ں���ˮ����������Ƭ���Ƕ���������7500�����ң��峤��135���ס����ٶ����ͷ���Ѿ��˻����㲿�ʸ�״�������������1��״�������Dz��������Ĺ����DZ����ģ���ر�����Ҫ��ؼ����������ɡ���ë�����������������У�ʹ��ˮ�����еĿ���������������ͨ��һ��ճĤ���Լ�������ȡӪ������ ˫�������嶯�������������Ҫ����ֳ�Ͳ��̶��������鱴�ȡ� ��2010��5�£�һ���й�˫�Ǹٵ��·������ڿ���Malacologia������������ƪ�����У�����ʹ���˶���ϵͳ����ѧ����Ѷ�����磺���ӷ���ѧ�����ʷ���ѧ�������̬ѧ���������ȷ�����ȥ�ѱ��ٵ��������·��ࡣ��������µķ��࣬˫�Ǹٵ����ֿ��Էֳ�324����Ч�Ŀƣ�������214����ֻ�л�ʯ������110���������������ڡ����������ΪWoRMS���ɡ� ������� ����� Eukarya �磺 ����� Animalia �ǽ磺 ����������ǽ� Eumetazoa ��δ�ּ��� ����Գƶ��� Bilateria ԭ�ڶ��� Protostomia ���ţ� ���ֶ������� Lophotrochozoa �ţ� ���嶯���� Mollusca �٣� ����� Scaphopoda Bronn, 1862 Ŀ �DZ�Ŀ �ܽDZ�Ŀ ����������嶯���ŵ�һ���١�ȫ���Ǻ�����ɳ��Ѩ�ӵ�һ��С�����嶯�����350�����ҡ�������ֱ��7000�ɼ����ټ��� ���dz�������������״����ϸ�䵶�ͣ����˿��ڡ��峤��14���ס����ִ��¶˿�����������Ͽ��ڳ����ォ���������ڳ������С�ͨ�����ֽ����壬��ʯ�������͵��ڲ����϶˿��ڱ����������Ӵ��� �������о��������ۣ������ֲ����ио�ϸ���� Ŀǰ�Բ��ܿ϶��������Ƿ�������༰���Ĥ�� ͷ����磺 ����� Animalia �ţ� ���嶯���� Mollusca �٣� ͷ��� Cephalopoda Cuvier, 1797 �Ǹ� �������Ǹ�(Nautiloidea) ��ʯ�Ǹ�(Ammonoidea) ���Ǹ�(Coleoidea) �����Ǹ�(Neocoleoidea) ͷ��٣�ѧ����Cephalopoda�������嶯���ŵ�һ���١���ʯ����һ�������ϣ��ֽ���786�֣���Ҫ�Ǹ������������㡣ͷ��ٶ���Ϊȫ����������ʳ�ԣ���������Գƣ���ͷ���㡢���������֡�ͷ�����������һ�Է�����ۡ���������ͷ�����ػ�Ϊ���©�����ʳ�ͷ���ࡣ©��λ��ͷ�����棬��ͷ������֮�䡣ԭʼ���������ǣ��ִ�����������ڿǻ��ǡ���Ϊ��״��һ�Ի���ԣ��Ķ���������Ŀ����һ�¡���ǻ�����Ƭ�ͳ��ࡣ��ϵͳ���У��йٷ��ѭ��ϵͳΪ�չ�ʽ��ֱ�ӷ����������̬���� ͷ��ٿɷ�Ϊ�������ĸ��Ǹ٣������ִ������Ǹ١�һ�������Ǹ٣�Coleoidea���ֳ�Ϊ�����Ǹ٣�����Ѿ���ʧ�����ڻ������Ǹٰ������㡢����������ȡ���һ�����������Ǹ٣�Nautiloidea���ֳ�Ϊ�����Ǹ٣������Ȼ���ڣ��˸ٰ��������ݵȡ�����������ľ�ʯ�Ǹ٣�Ammonoidea��Ҳ���Է���Ϊ�����Ǹ٣��ͼ�ʯ�Ǹ٣�Belemnoidea���� ͷ��ٷֲ������к����������ȣ�Ŀǰû�з�����Ӧ��ˮ�����࣬����Щ�ܹ���Ӧ��ͬ�ζȵ�ˮ�� ��ϵͳ����Ϊ ͷ�������ϵͳ�����������ϵͳ����Χ��ϵͳ�ͽ�����ϵͳ��ɣ��ṹ���ӡ� ͷ��ٱ���Ϊ����������������Ϊ�����и߶ȷ�չ��֪���ͽϴ���ԡ����ǵ��Աȸ���ٺ�˫�Ǹٶ����Ĵ���������֮�⣬ͷ��ٵı�Ƥӵ��һ�������ɫ��ϸ����chromatophore����ʹ�����ܹ����ɱ�ɫ�����й�ͨ��αװ�����о���ʾһֻ�����������������ĺ�ͯ���ܹ����п��ޡ����ľ�Ƚϸ��ӵĶ����� ����ͷ��ٵ���ϵͳ��������֮����Ϊ���ӵģ�������Ĥ��mantle�����Ӵ������ά��Ϊ������ѧ���õ�ʵ����ϡ� ͷ��ٵ��Ӿ�����ʵ��֤����ͨ�������ܹ���ʶ���ȡ���״����С��������Ĵ�ֱ��ˮƽ����ͷ��ٵ��۾����ܹ���Ӧ���ߵ�ƫ��ƽ�档���˾��ȵ��ǣ���Щ�ܹ���ɫ��ͷ��ٶ������ɫä�������ǽ���αװ��ʱ���ܹ����������������ı���������ɫ��ϸ���ı�Ƥ�������Ⱥͻ��ơ����ı���ɫ��ʱ��ʹ�õ��Dzʺ�ɫ��ϸ����iridophore���Ͱ�ɫ��ϸ����leucophores������Щϸ���ܹ���ӳ�����Ĺ��ߡ�ĿǰΪֹ��ֻ��һ������IJ�ɫ�Ӿ���color vision���õ�֤������Ϊө���ϣ�Sparkling Enope Squid���� ѭ��ϵͳ ͷ��������嶯�ﵱ��Ψһӵ�з��ʽѭ��ϵͳ�ķ��ࣺ����ӵ�������������е�Ѫ��������ѪҺ�����ģ�һ����������������������ͳ���Ѫ�ĵ�һϵͳ���ࣨsingle systemic heart���� �����������嶯��һ����ͷ�������һ�ֺ�ͭ���ӵ�Ѫ�൰�ף�hemocyanin���������������������������һ�㲸�鶯��ʹ��Ѫ���ء����ǵ�ѪҺȱ��ʱ����ɫ�����Ӵ�����֮��ʱ����ɫ�� �˶���ʽ ͷ��ٵ�һ���ж���ʽ���������䶯��������������ˮ����������Ĥ�е���֮��������ʹ�ռ���٣�����ˮ�������ݱ���ɵ�©���ܣ�hyponome�������ͨ���DZ�����ˮ����������ܹ���©�����Ʒ�������һ�������β���ƽ���Ϊ���ܵ��ƶ���ʽ�����Ч������������������ͣ���ҲʹһЩ���ྡ����ʹ�����ͱ����ƽ��� ��һЩ����������ܹ��ں������ߣ�ī����������ڶ�����Ĥ�ϵ���״�������ƶ��� ��ֳ���������� �����������⣬���Ǹٵ������ܶ��ҳɳ����٣���������µ����ֶ���������������������������������һ���������������͵�������hectocotylus������ֳ�ܣ�gonoduct������ĩ�ˡ����������������������������ԡ���һЩ����û�н���������ֱ�ӽ��ϳ��������������Ĥ�������ֱ�ӽ��䡣����������"���η�ֳ"��Ҳ����һ��ֻ����һ�Ρ�����һ����ĵ�֮��������� ���������Ǹ����С���η�ֳ�������������ϳ�����һ��ֻ�������ĵ��� �ݻ� ͷ����ں�������ڳ��֣��ڹ��������������ڼ�֧�䲢�ֻ���ˮ����̬�����������Ī�ؿ�����ͷ��ٵ�ԭʼ��̬��������������Ĵ���ȴ��������ţ�ĸ����������ں��������ڵ�ͷ���λ��ʳ�����Ķ��ˡ� �����ǹŴ��ļ�ʯ�ࣨBelemnoidea�������ִ�����ͷ���ࣨNeocoleoidea�������Ǿ�ʯ��ammonoids����������ӵ����ǵ�����������4.5����ǰ��3����ǰ�Ĺ������ֻ��������Ŵ���ͷ�������DZ�������Щ���ԭ����Բ״�����Ǻ��������������������������״�������ִ��������������Ȼ���ڿǣ��������ӵ����ǵ������ڰ��Ѽ;���ʧ�ˡ� ���� ͷ��� �������Ǹ٣�Nautiloidea�����ֳ������Ǹ� �̰���ʯĿ��Plectronocerida����ͷ����ں���͵����� ����˹ľ��ʯĿ(Ellesmerocerida)����������֮��ͷ��ٵ����� �ڽ�ʯĿ��Endocerida�� ���ʯĿ��Actinocerida�� ���̽�ʯĿ��Discosorida�� ��ֱ��ʯĿ��Pseudorthocerida�� ���ɽ�ʯĿ��Tarphycerida�� ������ʯĿ��Oncocerida�� ������Ŀ(Nautilida)�������ݺͻ�ʯ���ݣ������������� ����ʯĿ��Ascocerida�� ֱ��ʯĿ��Orthocerida�� ��ʯĿ��Bactritida���������������Ǹٺ;�ʯ�Ǹ����� ��ʯ�Ǹ٣�Ammonoidea�����������Ҳ�з�����Ѿ�ʯ���������Ǹ� ���ʯĿ��Goniatitida�� �ݾ�ʯĿ��Ceratitida�� ��ʯĿ��Ammonitida�� ���Ǹ٣�Coleoidea�� ��ʯĿ��Belemnoidea��������� ����ʯĿ��Aulacocerida�� �ռ�ʯĿ��Phragmoteuthida�� Hematitida ���ʯĿ��Belemnitida�� �����Ǹ٣�Neocoleoidea����Ҳ��Ϊ�����Ǹ� ʮ����Ŀ��Decapodiformes�� ������Ŀ��Spirulida����Ram's Horn Squid ī��Ŀ��Sepiida����ī�� ������Ŀ��Sepiolida��������������� ����Ŀ��Teuthida�������㡢���ϡ��������� ������Ŀ��Octopodiformes�� ��Ѫ������Ŀ��Vampyromorphida�����ʯ���ֳ������� ����Ŀ��Octopoda�������㣨Octopus�������٣�Argonauta����ˮĸ�٣�Amphitretus���� �������磺 ����� Animalia �ţ� ���嶯���� Mollusca �٣� ͷ��� Cephalopoda �Ǹ٣� �������Ǹ� Nautiloidea Ŀ�� ������Ŀ Nautilida ��Ŀ�� ��������Ŀ Nautilina Agassiz, 1847 �ƣ� �����ݿ� Nautilidae Blainville, 1825 ��/�� ����������(Allonautilus) ������������(Allonautilus perforatus) ��������(Allonautilus scrobiculatus) ��������(Nautilus) ����������(Nautilus belauensis) Nautilus clarkanus �ɿɶ�������(Nautilus cookanum) ����������(Nautilus macromphalus) ����������(Nautilus pompilius) Nautilus pompilius pompilius Nautilus pompilius suluensis ��������(Nautilus praepompilius) �װ������ݣ�խ�����ݣ�(Nautilus stenomphalus ) �����ݣ��������嶯�������ӡ�����̫ƽ�����������ձ��Ϸ�����������������������������쳼õȵ������з��֡�λ����������Ҫ���صķ����¿�������ǣ�������������Ϊ���յ���Ҫͼ�������������Ĵ�����֮һ�� �ִ���Ⱥ�����١��������������������ѹ��ˮѹ�������˹������� �������Ѿ��ڵ����Ͼ�������������ݱ䣬�����Ρ�ϰ�Եȱ仯��С�������������еġ��ʯ�������о��������������ѧ�ȷ����кܸߵļ�ֵ�� ���������ڵ�������������ݵ���Ϣ�ء� ����������ɫ��Ѥ�������ͻ����������Լ�ǿ�����ĿDZ����ᣬ���������̾����ǵı���ʰ�ɫ�������ɫ�������ƴӿǵ��겿���������ƽ��ϸ�ܣ���Ϊ���ɫ��������������ǹ⻬��Բ��״�����������죬�ʴ˵����������ݡ��� ���������������������ĵȽ����ߣ����Ƚ����߱�������ƽ�ָ������ܲ��ɷֵĹ�ϵ����ʹ�����������������֮�����������������Ȼ������ɫ�ʡ� ��������������ͨͷ��ٶ������ƣ���������ͷ���ʹ��롣�������ݵĴ�������ƫ�࣬���ɴ�90���������ݵĴ���û�����ܣ��֮�䲢�����ϵ����𣬶��ҿ����ջء������ݵij���ܴ���9�����ݣ��������� �����ڴ���ͷ��ٶ����У����������ݾ��б��ǣ�����ͷ��ٶ�����ޱ��ǣ����������������ȻҲ�пǣ�����ֻ��ĸ�����ڲ���ʱ�������ɫϵ������Ա����ѣ��������������ɷ��룬���Ѻ�㲻������ǣ�Ȼ�������ݵ������뱴���Dz������ġ� �����ݵı�������Ϊ26.8���֣�������������һ�㶼������20���֡����������ݣ�Nautilus macromphalus���������ݼ�����������С�����֣�һ��ֻ��16���֡� �����ݵĿDZ������ϵĸ���ָ�����ʮ��������Ŀ��ң������������ڵ����һ��������⣬���������壨��Ϊ���������������岻�ϳɳ�������Ҳ������������ƽ���������Ĥ�������̼������л����ʣ�������һ��ո�µĸ��塣������֮����һ���ᴩ��ϸ�ܣ����������������������֮�У�ͨ������ĵ��ڣ��������帡�������С��������������������ṹҲΪ���ཨ��DZͧ�ṩ����У�������ҵ�DZͧҲ�ԡ������ݡ������� ������Ϊůˮ�Զ����������ˮ��Ϊ19-20�棬һ��������50��300����ĺ����У�ͨ��ҹ���Ծ�����Ե��ܵ�Сз��СϺ�ȼ��ද������Ϊʳ���ռ������ɺ����dz�����ҷ��У��Դ���������ʯ��ЪϢ�����������������������ѿǶ���û����տ���������Ư�ƣ��о���Ư��·�߶������ķ�����һ�����塣 �����ݴ������壬��������������ÿ�����һ�Σ�һ�㽫�Ѳ���dzˮ��ʯ�ϣ�������12���£��³��ǵ�С������һ���峤Լ3���֡� ����������һ��Ϊ��ʮ�꣬��ͷ��ٶ����������ϳ��Ķ�� ���ƺ������ʹ�� �����ݺţ��ƻ�С˵�����ա������ɵĴ�����֮һ��������������鹹��DZͧ�� �����ݺţ������ϵ�һ��ʵ���������۵ĺ˶���DZͧ�� �����ݣ�������ɻ����д������� ��ʯ�Ǹ��磺 ����� Animalia �ţ� ���嶯���� Mollusca �٣� ͷ��� Cephalopoda �Ǹ٣� ��ʯ�Ǹ� Ammonoidea Zittel, 1884 Ŀ ��ʯĿ Ammonitida ���ʯĿ Goniatitida �ݾ�ʯĿ Ceratitida ��ʯ�Ǹ٣�ѧ����Ammonoidea����һȺ�Ѿ�����ĺ��������ܳƣ��dz��ʺ���Ϊ����ʯ������ѧ�ҿ���ʹ��������ȷ�����о�ʯ��ʯ�ĵز���������ʯ�Ǹ����ִ��ͷ��ٹ�ϵ��ӽ��Ŀ��������Ǹ٣��������������Ǹٵ�������Ŀ�� ��ʯ�Ǹ�Լ��־��������������ͳ��ڵ�һ�γ����ڵ����ϣ���������һ���ڰ��Ѽ���������� ����ѧ�Ҹ��ݻ�ʯ�Ŀ��ҵĽṹ����ǵĻ��ƣ�Ŀǰ����ʯ�Ǹٷֳ�3��Ŀ����ʯĿ�����ʯĿ��ݾ�ʯĿ�����о�ʯĿ�ֳ������֪����Ŀ�� ��ʯ�Ŀ���ƽ�����������״������Գƣ��DZ���⻬���ϸ���������ƣ���Щ����������Σ������⡢���ߡ����ʹ̵ȡ� ��ʯһ��Ư���ں�ˮ�ϲ㣬���澭���Ǽ���ȱ������û�������ʯ���������ף�����û��ϸ���ֽ�����ʱ�Ѹ���ˮ�����ʱ仯�����Ϳ������ܽ�ȣ������������κ�̼���Ρ���ʯ��ʯ������һȦһȦ�Ŀ����ʣ���˱���ܶ���ʱ걾�� ���ʯĿ�磺 ����� Animalia �ţ� ���嶯���� Mollusca �٣� ͷ��� Cephalopoda �Ǹ٣� ��ʯ�Ǹ� Ammonoidea Ŀ�� ���ʯĿ Goniatitida Hyatt, 1884 ��Ŀ ���ʯ��Ŀ ԲҶ��ʯ��Ŀ Tornoceratina Anarcestina Clymeniina ���ʯĿ��Goniatitida����һȺ�Ѿ�����ľ�ʯ�Ǹ�����������������ϵ�Ͻӽ����������ݹ�ϵ��Զ�����ʯĿ�����������������ڣ���Լ4����ǰ������ʢ��ʯ̿��������ͣ�����ڹ�����ĩ�ڣ�Լ2��4500����ǰ����� ���е����ʯ�ӵ����ǣ�Ӳ���ڲ�������ָ��ɶ����Ŀ��ң���������ܶ����������塣�ֵ����ʯ��ֱ����С��15���֣���������С��5���֡����ʯ������ͨ������������״�Ļ��ƣ����������ľ�ʯ�ͬ���������ľ�ʯ����Ǵ�ֻ�в����������ƣ���Щ��������û���������ơ� ��ʯĿ�磺 ����� Animalia �ţ� ���嶯���� Mollusca �٣� ͷ��� Cephalopoda �Ǹ٣� ��ʯ�Ǹ� Ammonoidea Ŀ�� ��ʯĿ Ammonitida Hyatt, 1889 ��Ŀ ��ʯ��Ŀ Ammonitina ����ʯ��Ŀ Ancyloceratina Ҷ��ʯ��Ŀ Phylloceratina �ھ�ʯ��Ŀ Lytoceratina ��ʯĿ��ѧ��Ammonitida����һ���������ͷ���ࡣ�����Ƿdz��õ�ָ��ʯ�����������������ڵĵز�������������ʯĿ�Ľ��ײ����ֽ�������Ƶ������ݣ������ǰ�צ�㡢��������������Ǹ١� ��ʯĿ�Ļ�ʯ��һ���ƽ��״����Ҳ�гʴ�ֱ����״��������״����������ƾ�ʯĿΪ�����ɵĽǡ�������ǵ���״���������������ǣ���ѧ�������Դ���ȡ�ġ����µ��������ϣ���ġ��ǡ����ָ�����β�� ��ʯĿ�����������ĸ���Ŀ���ֱ�Ϊ��ʯ��Ŀ����٪�����ϰ��Ѽͣ�������ʯ��Ŀ����٪�����ϰ��Ѽͣ���Ҷ��ʯ��Ŀ�������������ϰ��Ѽͣ����ھ�ʯ��Ŀ����٪�����ϰ��Ѽͣ�����������������ѧ�۴ԡ���The Treatise on Invertebrate Paleontology��ȴֻ�����˾�ʯ��Ŀ���ھ�ʯ��Ŀ��Ҷ��ʯ��Ŀ������֮�����ھ�ʯ�Ǹ�֮�£�������ʯ��Ŀ��Ϊ�ھ�ʯ��Ŀ�µ�һ�����ơ�������ѧ�߽���������Ŀ�����ھ�ʯĿ�С� �����۴���������ʯ��Ŀ����٪�ͳ�����Ҷ��ʯ��Ŀ���ھ�ʯ��Ŀ���������� ��ʯĿ��������������������岿������Ϊϡ�С����о����ǻ��ھ�ʯĿ�ĿǼ��ǵ�ģ�͡� �־�ʯĿ������������Զ�ź���Ŀ���ˮ���Ǻ��ס������������ǵĻ�ʯ����������û�е��ܶ���ĵز㷢�֡��ܶ��ʯĿ���Ŷ�����Ӿ���֣��������ʯ�������ǵĿDZ�ƽ��������״�������Σ���Ҳ��һЩ���������λ���ܵġ���ʯĿ�����������ܻ���ʳ���ࡢ���༰����ϸС������ǵ���а���ˮ�������࣬������ƣ��ڻ�ʯ���ľ�ʯĿ����һЩ���ݺۼ������ǿ��ܻ���ī֭���ӱ���ʳ����ΪһЩ��ʯ�걾��������Щī֭�� ��ʯĿ�����岿��ռ���˿�ĩ�����ڡ������ϸС�Ľڶ��м�������������������ά�ָ������ʴ˽�ϸС�IJ��ֻḡ�ڽϴ��֮�ϡ� ��ʯĿ���м���IJ��ֳ�Ϊ����������������ס�ң�ס��֮����һ���ܱ��ļ����������ʯĿ����ֻס���������ס���С���������ʱ�����ڿ��ڴ����ӽ��¼��ϴ��ס�ҡ�����ϸ���ɾ�ʯĿ�����壬������������п��õ�ס�ҡ����������ĸ����������ã���ʯĿ���Խ���Щס�ҵ�ˮ�ݳ��ߣ��Ӷ����ƿǵĸ�����ˮ���������³��� ��ʯĿ������������Ҫ��֮ͬ��������������ϸ�����ظ��洩������ģ����������������λ�ڽӽ�����λ�á� �ֽ������ݿǵĴ�С�����Ա��йأ����ԵĿǽ�ϸС�������ԵĿǽϴ������������ο���Ҳ�ھ�ʯĿ�г��֣������ý��������Ǵ�С�ı��졣������Ҫ�ϴ�Ŀǿ�����Ϊ����֮�á�٪������ŷ�ĶԾ�ʯ��������һ�����ӡ���ʯĿ�ǵ���������ֻ�ǽ���ķ��֡��������ִ�С�IJ��챻��Ϊ�ǻ�Ϊ���IJ�ͬ���֡� �־�ʯĿ�Ƕ��dzʱ�ƽ��״����Ҳ��һЩ�ǽӽ���ֱ�ģ���˾�ʯ��������ٿǵ�������������ʯ����˹�ؾ�ʯ��������չ��������������ʱ�DZ�ֱ�ģ������ʯ�������ּ���ȫչ������̬�ڰ��Ѽͳ��ڿ�ʼ��ö�������������Ϊ�����ξ�ʯ���� ���ξ�ʯ�������˼��쳣�������ձ���ʯ�����Ǿ�����һ�Ų������������û�����ԶԳƵľ���������������ʱ���Ϳ��Լ����ձ���ʯ������U�ε���ά���硣 ��ʯĿ�ǵı����Ұ�ܴ�һЩ���������������ͺܹ⻬���������ֽ�������ݣ�һЩ����������������߹ǣ������м����������ľ�ʯĿ�����ѼͽϺ��ڳ��֡� һЩ��ʯĿ����ͬһ���״���һ�Է���ʯ��һͬ�����֡�������Ϊ��Щ��������������ǣ�����������һ������������ȴ��Ϊ����ʵ��������Щ��ͳ��Ϊ�ڸǣ���һ��ij�Ϊ���ڸǣ��ɶԵ�����˫�������ǵ��ڸǻ�˫�������ܱ������������Ҷ�����Ƿ��롣���ڸ�ͨ��������������������Ѽ͵ľ�ʯĿ����˫������������������ľ�ʯĿ��˫����ǶԳƵģ���С�����һ���� �ڸ��ж�����״�����������ƿ�Ҳ���в�ͬ�����ǣ��������Ǻ�ϡ�У��������Բ������һ��ڸ���������һ�����֣�һЩ�ڸ���������Ϊ���������� ��Ȼ��ʯĿ�Ļ�ʯ�����ܶ࣬��������ȴʮ��ϡ�١�����һЩī֭�Ҽ����ܵ����������⣬��û�з����κ��������֡����������кܶഥ��������� ٪���缰���ڵľ�ʯĿ���ٻᳬԽֱ��23���Ĵ�С������٪�����ڼ����Ѽ����ڣ��Ϳ�ʼ������һЩ�ϴ�����壬��Ӣ�����ϲ���̩̹��ʯֱ���ʹ�53���ף������Ѽ͵¹���Parapuzosia seppenradensis���д�2��ֱ������������֪���ľ�ʯĿ����Parapuzosia bradyi��ֱ����1.37�ס����⣬��Ӣ�����ױ��Ƿ�����ֱ����2.3��δȷ���֣�һ�����ȷ�ϣ�������֪���ľ�ʯĿ�� ���ھ�ʯĿ��������Ӿ���εģ���������������Ϣ�ں������ϵ�ˮ�����������ͻ��³������������ڳ����£�ʬ��ᱻϸ����������ά��ƽ�⡣����Ȼ�أ��仯ʯ�ܱ߾ͻ���ֿ����ʵ�ͬ�ij�������Ϊ��ˣ�Ҳ�ǿ��Ժúñ����ʯĿ��ʯ��ԭ�� ������з��ֵľ�ʯĿ���涼�ᱣ������ĸ���Ϳ�㣬��Ӣ�����Ŀ���ʯ�� ���������ô�İ��Ѽ�Ƥ��������ҳ��������ľ�ʯĿ��ʯ�������˾�ʯ������ʯ������ʯ��װ�״���ʯ��Jeletzkytes���뼰����չ������̬����Щ��ʯ�ֶ��ܱ���ԭ�еĿǡ� ������ʯ�����������˹�Ӽ����ô���ʡ���ֵģ������к�ɫ����Щ��ɫ��ʯĿ�ڴ�ĥ���б�ʯ�����ء��������ֺ�ɫ�ڻ������ľ�ʯĿ�������ڣ�����������϶��˶����һ�㡣 ���������Ժ�ʯĿ��ʯ�Ϳ�ʼ���٣�ֱ������������ȫ��ʧ����������������6500����ǰ�İ��Ѽ�-����������¼����ڰ��Ѽ����ڵ����OȺ������û�о�ʯĿ���棬��ȴ����巢��ѧ�ķ�Ұ��������һЩ�������ࡣ����ʯĿ������������ײ�����ܱߵĸ��������������ʧ����ɾ�ʯĿ��ֳ�����ѡ� �������͵�ŷ�ޣ���ʯĿ��ʯ����Ϊ��ʯ���˵��ߣ����ֱ���Ϊ��ʯ���ഫ�����ɻ����ص�ϣ���Hilda of Whitby����ʥ������˵�ʥ���¼���֤�ݡ����˻��ھ�ʯĿ�Ŀ��ڿ����ߵ��档��ʯĿ���Ჴ����������ӣ�Gandaki river������Ϊ���������ķ����ӡ�Ƚ�������������ʪū�����֡� ����ʯ��Ŀ�磺 ����� Animalia �ţ� ���嶯���� Mollusca �٣� ͷ��� Cephalopoda �Ǹ٣� ��ʯ�Ǹ� Ammonoidea Ŀ�� ��ʯĿ Ammonitida ��Ŀ�� ����ʯ��Ŀ Ancyloceratina Wiedmann, 1966 ���� Ancylocerataceae Douvilleicerataceae ����ʯ���� Turrilitaceae ��ɳҮ˹��ʯ���� Deshayesitaceae Scaphitaceae ����ʯ��Ŀ��ѧ��Ancyloceratina����һ��������ľ�ʯ����ʯĿ�³ھ�ʯ��Ŀ�Ľ��ס�������٪�����ڿ�ʼ�ݻ��������Ѽ��ڼ�dz��ձ鼰�����������DZ鲼�����ޣ�������Ϊָ��ʯ���ֹ���ʯ��Ŀ�ĿǶ���������״�����������ξ�ʯ�� ��ԭʼ�Ĺ���ʯ��Ŀ����״�ӽ�������������ֻ������ĩ����˿��ţ��������µ�һ��������������״���ԣ������IJ��־��Ǹ���IJ��֣���������ס�ҡ����Ӱ�������ʯ��Protanisoceras��Tropaeum���Ϻ��ڵ���̬��֮������IJ������Ptychoceras�ĿǾ��������ĸ�����һ�������ӵ�Ͳ������ʯ����״������һ�������롣 ���˰��Ѽ����ڣ����ǵ���״��Ϊ��䣬��������ȫ��ֱ�ĸ˾�ʯ��Sciponoceras��Ťת����״������ʯ�������ι�״���ձ���ʯ��һЩ��̬���ۺ��˶����״������Nostoceras����ʼ��������ʯ�������Σ���������ʯ��ˮƽ�Ĺ��� �������й���ʯ��Ŀ����״���������ġ��ܶ����ڵ���̬�����������ձ�ľ�ʯĿ�ֱ������һЩչ���ķ�֧����Ҳȡ����������״���������İ�������ʯ��װ�״���ʯ���������� ����ʯ��Ŀ����̬����������һ����Կ϶�����չ����̬�IJ����ʺ���Ӿ��һЩӵ�п���ʽ�ǵĺ����������У������Ҳ�Ǹ�������֮�ϣ�����Ӿ��ɼ�����谭�����ԣ����Ǻ��п���ֻ�Ǹ��εģ��Գ���������ϸС����ֻ��ǵ��ܵġ� ����ʯ��Ŀ�Ĵ�С����ܴ���ϸС��Ptychocerasֻ��Լ3���׳������˾�ʯ��Diplomoceras�Ϳ��Գ���1-2�ס�һЩ�����繳��ʯ�㷺�ֲ���ŷ�Ǵ�½���������ϼ��ޣ�����һЩ�Ϳ���ֻ����������½������ ��ʯĿ�磺 ����� Animalia �ţ� ���嶯���� Mollusca �٣� ͷ��� Cephalopoda �Ǹ٣� ���Ǹ� Coleoidea �¸٣� ��ʯ�¸� Belemnoidea Ŀ�� ��ʯĿ Belemnitida Zittel, 1895 ��ʯĿ��ѧ����Belemnitida����һ���Ѿ������ͷ����������������������Ѽ�֮�䡣��ʯ������涼���ִ��������൱�ӽ����������ִ���������ϵ���С���ʯҲӵ��ī֭������������10����Լ��ͬ���ȵĴ������в�û���رĴ��� ��ʯ��٪������Ѽ�ʱ�����൱�࣬�������ǵĻ�ʯ���������ĺ����Ҳ����൱�ḻ�����������ž�ʯ����֡�������ʯҲ����ʯһ���ڰ��Ѽ�ĩ������ˡ� ��ʱ��ʯĿֻ��������������Ŀ�� Belemnitina Belemnopseina Belemnotheutina ����������Ŀԭ�����ڼ�ʯĿ����ʱ�ѷ����������ࣺ Aulacocerida Phragmoteuthida Diplobelida ���Ǹ��磺 ����� Animalia �ţ� ���嶯���� Mollusca �٣� ͷ��� Cephalopoda �Ǹ٣� ���Ǹ� Coleoidea Bather, 1888 �¸ټ�Ŀ ��ʯ�¸� Belemnoidea Aulacocerida �ռ�ʯĿ Phragmoteuthida Donovaniconida ��ʯĿ Belemnitida Hematitida �����¸� Neocoleoidea Boletzkyida ����Ŀ Sepiida ������Ŀ Sepiolida ������Ŀ Spirulida ǹ��Ŀ Teuthida ����Ŀ Octopoda ������Ŀ Vampyromorphida ���Ǹ٣�ѧ��Coleoidea����ͷ����µ�һ�����嶯����Dz����������Ǹٰ���Ӳ�ǣ����ֻ���������Ƹ������ڹǡ�һЩ����������ȫû�й�ͷ��һЩ����������֧�š� ���Ǹٵķ������Դ��ֵ��������ṹ������ġ����������ԭʼ�ļ�ʯ�¸���������10ֻ���֡����ִ�����������һ�Դ��ֿ����Ѿ��������˻��ˡ�ʮ����Ŀ�ĵ��Ķ��ֱ۽�����Ϊ�����̵ij����֡�������Ŀ�ĵڶ����ֱ��в�ͬ�ĸı䣬��������Ŀ�ĸ�Ӧ˿״������Ŀ������ȫ�˻��� �ݻ���ʷ ����ϵ����Ǹ���Դ��Լ��3��3000����ǰʯ̿�͵��������ȼ͡�Ҳ�з���һЩ��������͵Ļ�ʯ������ѧ�߲���ͬ���������Ǹ١� ����ʯ̿�ͣ����Ǹ��Ѿ��ж�������̬����Ȼ��Щ���ͳ�϶�����Ϊ��ʯ�¸٣������ǵķֱ�������ʾ���Dz��ǽ��ס� ���� ͷ��� �������Ǹ� ��ʯ�Ǹ� ���Ǹ� ��ʯ�¸� ʼ������ Hematitida �ռ�ʯĿ Donovaniconida Aulacocerida ��ʯĿ �����¸� ʮ����Ŀ Boletzkyida ������Ŀ ����Ŀ ������Ŀ ǹ��Ŀ ������Ŀ ������Ŀ ����Ŀ ����Ŀ ʮ����Ŀ�磺 ����� Animalia �ţ� ���嶯���� Mollusca �٣� ͷ��� Cephalopoda �Ǹ٣� ���Ǹ� Coleoidea ��Ŀ�� ʮ����Ŀ Decapodiformes Ŀ Boletzkyida ������Ŀ(Spirulida) ī��Ŀ(Sepiida) ������Ŀ(Sepiolida) ����Ŀ(Teuthida) ���� Decembrachiata Winckworth, 1932 ʮ����Ŀ��ѧ����Decapodiformes����ͷ������Ǹ������¸�֮�µ�һ�����嶯������¸ٵ���һ������ǰ�����Ŀ�� �����Ǹٵ���������һ����ʮ����Ŀ������ͬ����10ֻ���֣�������ĶԴ����ݱ��Ϊ�������̵ij����롣 ���� ʮ����Ŀ (Decapodiformes) Boletzkyida ������Ŀ(Spirulida)��Ram's Horn Squid ī��Ŀ(Sepiida)��ī�� ������Ŀ(Sepiolida)������������� ����Ŀ(Teuthida)�����㡢���ϡ��������� ������Ŀ�磺 ����� Animalia �ţ� ���嶯���� Mollusca �٣� ͷ��� Cephalopoda �Ǹ٣� ���Ǹ� Coleoidea ��Ŀ�� ������Ŀ Octopodiformes Berthold & Engeser, 1987 Ŀ ����Ŀ Octopoda ����Ŀ Trachyteuthis ������Ŀ Vampyromorphida ���� Octobrachia Fiorini, 1981 Vampyropoda Boletzky, 1992 ������Ŀ��ѧ����Octopodiformes����ͷ������Ǹ������¸�֮�µ�һ�����嶯������¸ٵ���һ�������ʮ����Ŀ�� �����Ǹٵ���������һ����������Ŀ������ͬ����10ֻ���֣�����ͬ���ֵĵڶ����ֱ��в�ͬ�ĸı䣬��������Ŀ���ݱ��Ϊ��Ӧ˿״�������Ŀ������ȫ�˻���  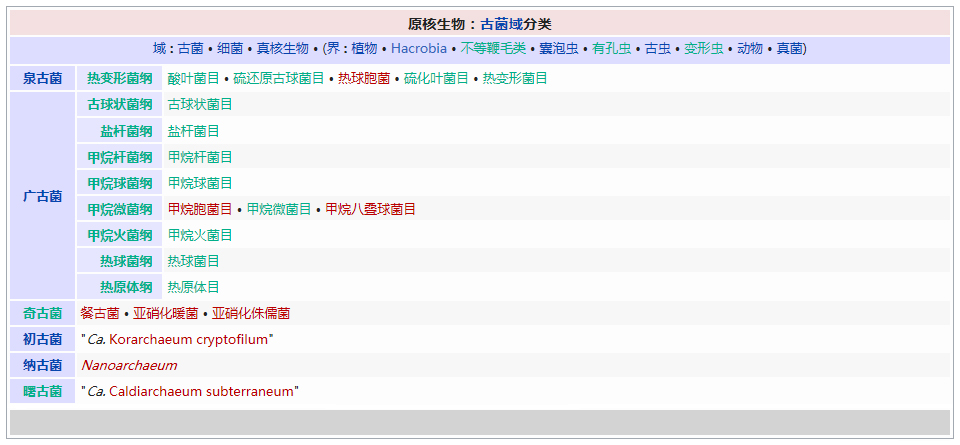    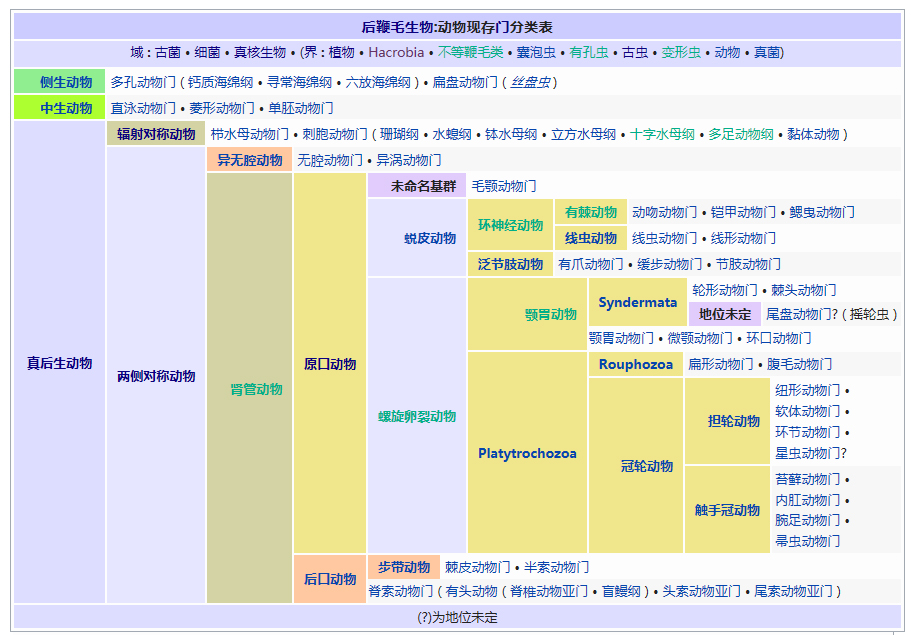      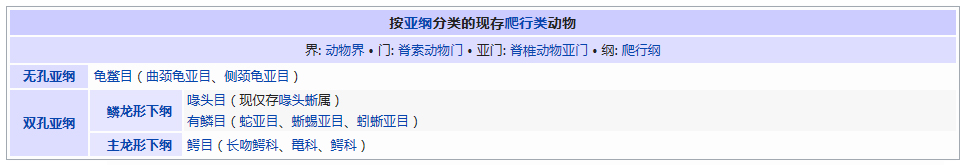 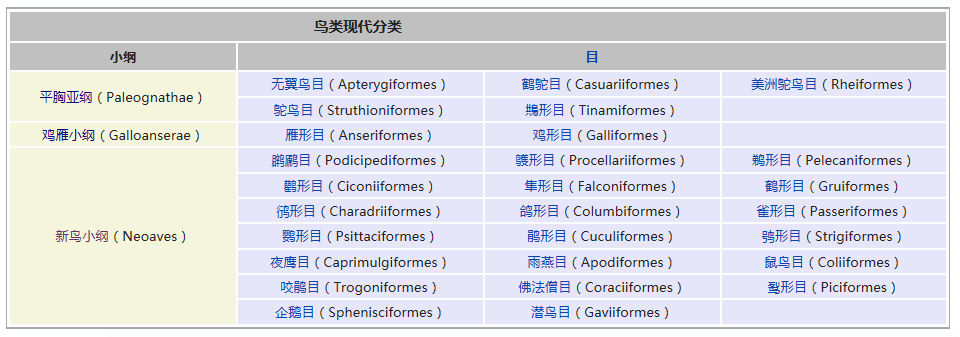 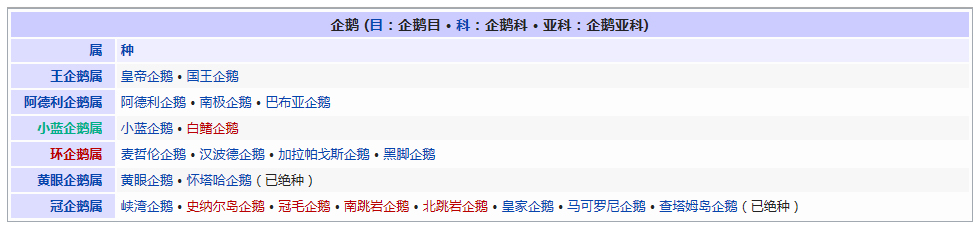 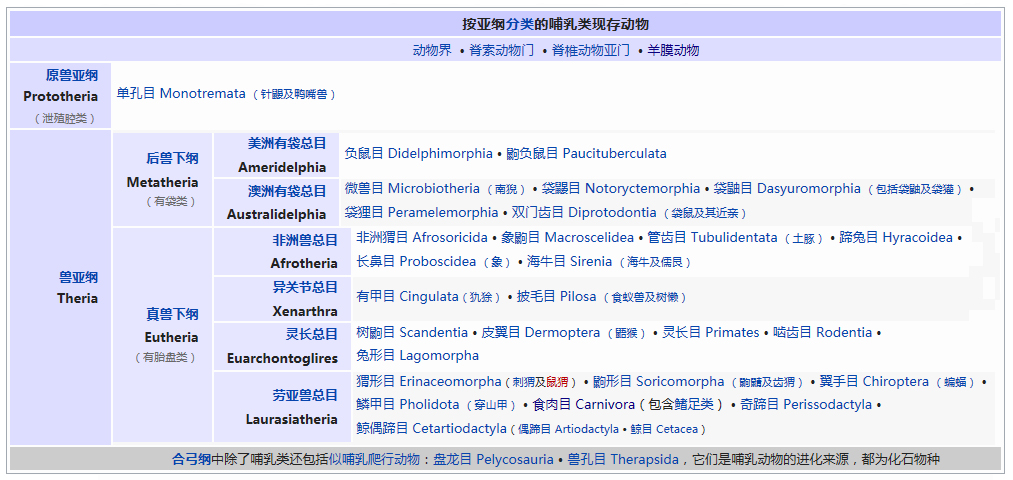    |
| ������ͣ� | |
| W(��15����Ϣ) |
|
| �����Ϣ�� | |
| �������� Э������ʱ ������ϵ������ ǰ����� ���ǹ��� | ������ ������ �Ǽ����� ��С���Ǵ� ������ |